

Longueur moyenne bras étendus inférieure à un mètre
Bras quatre fois plus longs que le manteau
Ocelles pourpres à brun foncé cerclés de clair et bordés de brun, à la base des paires de bras II et III
Livrée la plus commune : brun rouge avec ou sans taches blanches
Grande variabilité dans l'apparence (chromatique et texturale)
Poulpe diurne, gros poulpe bleu (FAO), zourite (La Réunion, Seychelles), ourite (Maurice, Rodrigues), ces deux derniers noms étant dérivés du mot malgache [orita], désignant le poulpe.
Common reef octopus, big blue octopus, day octopus, daytime octopus, Cyane's octopus (GB), Polpo diurno, polpo di barriera (I), Pulpo azulon (E), Gemeiner Riff-Krake (D), Gewone rif-octopus (NL), Seekat (Afrique du Sud), Orita (Madagascar), Day squid, he'e (Hawaï), Eke Pae, mangamanga (Cook), Fe'e (Samoa)
Octopus marmoratus Hoyle, 1885
Octopus horsti Joubin, 1898
Octopus herdmani Hoyle, 1904
Octopus glaber Wülker, 1920
Octopus cyanea var. gracilis Robson, 1929
Callistoctopus magnocellatus Taki, 1964
Mer Rouge et Indo-Pacifique
Zones DORIS : ● Indo-Pacifique, ○ [Mer Rouge]Ce poulpe est présent en mer Rouge et dans tout le domaine indo-pacifique de l'Afrique du Sud jusqu'à Hawaï (dont Mayotte et La Réunion, la Nouvelle-Calédonie et la Polynésie). Il fréquente les zones tropicales et subtropicales.
L'espèce est benthique*, on peut la trouver jusqu'à 60 mètres de profondeur. On la rencontre sur des types de substrats* très divers, sableux, sablo-vaseux, sablo-détritique*, rocheux, dans les herbiers, etc. mais toujours en milieu corallien ou proche d'un environnement de ce type.
Octopus cyanea est un poulpe de taille moyenne. Son manteau* peut mesurer 16 cm et les bras 80 cm. La mesure du manteau est dorsale, elle va du milieu de l'espace interorbital à la limite postérieure du corps (N.B : les tailles données pour les poulpes dans les publications scientifiques sont celles du manteau). Cependant, des individus plus grands ont été observés.
Tous ses tissus sont mous à l'exception des pièces cornées qui arment le bulbe buccal, et de la radula* qu'il contient.
Le manteau (ou complexe palléo-viscéral) a la forme d'une poche oblongue. Il est largement fendu en partie antérieure d'un flanc à l'autre en passant par la face ventrale (fente palléale*). Cette fente laisse apparaître des deux côtés la cavité* palléale, espace interne délimité par les lobes du manteau. Un entonnoir orientable (aussi appelé siphon* ou hyponome*) en forme de cône est situé dans cette cavité en partie antéro-ventrale et la dépasse latéralement de l'un ou l'autre côté de l'animal.
Tête et bras forment le céphalopodium. La tête se signale essentiellement par deux grosses protubérances globuleuses qui abritent les yeux, situés latéralement. Elles sont susceptibles d'être érigées (surveillance, alerte, curiosité), ou au contraire rétractées (stress, camouflage). Elles peuvent aussi être surmontées de cirrhes* en forme de cornes dans les livrées de stress. Une large zone circulaire entoure les yeux, elle est ornée par deux séries alternées de cinq bandes, les unes foncées et les autres claires, qui rayonnent à partir de l'œil. Cette caractéristique est présente dans la plupart des livrées, mais elle est peu discernable dans les livrées sombres, et est parfois réduite à la partie horizontale des motifs foncés dans les livrées claires. Les yeux ont une pupille horizontale. Le bulbe buccal, dont les pièces cornées ont la morphologie d'un bec de perroquet, est situé ventralement au centre de la couronne formée par la membrane brachiale (la membrane entre les bras).
Les bras sont puissants et de taille approximativement égale, la quatrième paire étant légèrement plus longue que les autres. Ils sont environ quatre fois plus longs que le manteau et sont armés de deux rangées de ventouses. On en compte entre 180 et 230 sur le bras hectocotyle* du mâle, et entre 400 et 500 sur les autres bras. Un ocelle* ovale, pourpre à brun très foncé entouré d'un anneau clair bordé de brun, se trouve sur la membrane brachiale entre les yeux et la base des paires de bras II et III : ces « faux yeux », qui sont une caractéristique de l'espèce, ne sont pas fréquemment visibles dans la mesure où l'animal peut les faire disparaître à volonté.
La livrée est extrêmement variable. La couleur ordinaire de l'animal au repos est brun rouge. Elle est souvent parsemée de taches claires plus ou moins diffuses sur le manteau, des taches blanches, rondes et de taille décroissante étant alignées sur les bras en séries longitudinales. La texture du tégument* est alors lisse à légèrement grumeleuse, les sillons entre ces reliefs dessinent une fine réticulation plus foncée. L'intérieur de la cavité palléale, visible quand l'animal y aspire l'eau pour respirer, est blanchâtre. L'entonnoir servant à expulser l'eau est beige à ocre ou orange foncé.
L'un des premiers signes de stress sur cette livrée est l'apparition d'une longue bande médiane blanche partant de la jonction de la première paire de bras et s'achevant à la pointe postérieure du manteau en s'élargissant entre les yeux. Pour l'ensemble des variations de livrée, voir le paragraphe « Variations de l'apparence » en Divers Biologie.
La capacité des poulpes à changer d'apparence rend particulièrement difficile l'identification in situ des espèces quand les principaux paramètres morphologiques sont sensiblement identiques (taille maximale, longueur des bras relativement à celle du manteau et différences de longueur entre les paires de bras notamment). Cela d'autant plus que le signe caractéristique d'O. cyanea, les faux yeux présents à la base des paires de bras II et III, est rarement manifesté par l'espèce. Le meilleur moyen pour l'identifier reste le fait qu'O. cyanea est un des rares poulpes à être diurne, ce qui limite beaucoup le choix. La distribution, généralement utile pour resserrer un champ d'identification, est en ce qui concerne ces espèces assez aléatoire puisque les spécialistes en débattent encore pour beaucoup d'entre elles. Pour mentionner quelques uns des fauteurs de confusion possibles :
Octopus vulgaris : son apparence, ses comportements et ses habitudes sont très proches de ceux d'O. cyanea, mais il ne présente pas les motifs rayonnants autour des yeux fréquents chez ce dernier. Il peut pourtant avoir deux marques horizontales sombres de part et d'autre de chaque œil, notamment en livrée claire, ce qu'on peut trouver aussi chez O. cyanea dans le même type de livrée. Cette espèce est diurne et nocturne, bien qu'elle semble essentiellement nocturne en Méditerranée. Sa distribution stricte la signale en Méditerranée et en Atlantique Est. Cependant, s'agissant du « complexe » O. vulgaris (un complexe signifie qu'il pourrait y avoir plusieurs espèces cryptiques* dans le groupe), la distribution s'étend à toutes les zones tropicales, subtropicales et tempérées de la planète. Une étude de 2010 se fondant sur des données génétiques montre de surcroît que O. vulgaris sensu stricto (autrement dit il s'agit bien de l'espèce décrite par Cuvier) est présent aux îles Saint-Paul et Amsterdam, dans le sud de l'océan Indien. Son signalement a aussi été confirmé des côtes indiennes en 2011 et il pourrait donc se rencontrer dans d'autres zones de l'océan Indien.
Callistoctopus macropus : son manteau est plus petit que celui d'Octopus cyanea et sa livrée généralement rouge à points blancs peut tromper, mais ses bras sont nettement plus longs et plus fins, et il est nocturne (bien qu'il ait montré une activité diurne en laboratoire). Il est supposé avoir une distribution circumglobale*, ce qui signifie qu'on peut le trouver dans toutes les eaux chaudes et tempérées du globe.
Callistoctopus luteus : il est un peu plus petit qu'Octopus cyanea (longueur moyenne du manteau 10 cm), sa livrée ordinaire est rouge foncé à points blancs mais ses bras sont très longs (jusqu'à 6 fois la longueur du manteau), et les bras dorsaux sont plus longs que les bras ventraux (ils sont sensiblement égaux chez O. cyanea). Il est nocturne. On le trouve dans le nord et l'ouest du Pacifique, mais son signalement a été confirmé par des analyses ADN sur les côtes indiennes en 2011.
Callistoctopus ornatus : à nouveau une livrée ordinaire rouge à points blancs, mais elle présente généralement des lignes longitudinales blanches à crème sur le manteau. Ses bras sont beaucoup plus longs que ceux d'Octopus cyanea (leur portée peut atteindre 2 mètres pour un manteau de 13 cm) et les bras dorsaux sont plus longs et plus fins que les autres. Il est nocturne et se rencontre dans l'océan Indien et l'ouest du Pacifique (mais certaines distributions le mentionnent jusqu'à Hawaï et l'île de Pâques).
Octopus cyanea se nourrit de crustacés (avec une préférence pour les crabes) et de poissons. Il peut aussi chasser de petites murènes. Contrairement à de nombreux autres poulpes, il ne semble pas consommer de gastéropodes à coquille ni de bivalves. Diurne, il se nourrit cependant de préférence au lever et au coucher du soleil. A l'instar des autres poulpes, il est carnivore dès l'éclosion et occupe une position élevée dans le réseau trophique*.
Méthodes de chasse : il y en a deux.
Mâles et femelles :
Les mâles sont sexuellement matures 6 à 7 mois après l'installation* sur un récif. Leur maturité sexuelle ne dépend pas de la taille mais de l'âge : elle peut survenir chez des individus pesant entre 200 gr et 6 kg, la vitesse de croissance des juvéniles dépendant de l'abondance de la nourriture dans leur environnement. Il n'y a pas de dimorphisme* (hors parfois une différence de taille) ni de dichromatisme* sexuels. Sauf exceptions liées au fait que les phéromones* d'une femelle ont attiré deux mâles, il n'y a pas non plus de rivalité, ni de combats de mâles pour la possession des femelles. L'accouplement semble se faire non sélectivement au hasard des rencontres. Les œufs des femelles sont encore immatures juste avant la ponte (les femelles ont alors 10 à 12 mois), mais elles peuvent recevoir et stocker des spermatozoïdes* issus de nombreux mâles différents bien avant cet âge : les deux sexes s'accouplent avec des partenaires différents à partir de l'âge de 4 mois. L'espèce se reproduit en toute saison de l'année avec un pic de ponte au début de la saison chaude, et un pic secondaire moins important au milieu de la saison froide.
L'accouplement :
La cour est très limitée : le mâle s'approche et touche plusieurs fois le corps de la femelle du bout d'un bras. Ces premiers contacts semblent destinés à confirmer le sexe du partenaire, qui ne paraît pas reconnu comme tel visuellement mais pourrait l'être chimiquement grâce aux chémorécepteurs* contenus dans les ventouses. L'accouplement commence par une lente approche du mâle, en livrée bordeaux foncé à marques claires, près d'une femelle généralement en livrée de camouflage.
Chez le mâle, le bras droit de la troisième paire est tenu en position haute et il est enroulé sur lui-même. Il s'agit du bras hectocotyle*, autrement dit d'un bras modifié de façon à pouvoir transférer des spermatophores*, les gamètes* étant produits par des gonades* situées dans la partie postérieure de l'organisme, en arrière de l'appareil digestif. Restant à distance de la femelle, et si elle est réceptive, le mâle déroule lentement son bras hectocotyle pour en introduire l'extrémité dans la cavité palléale de la femelle, puis dans son oviducte, et y déposer les spermatophores qui s'y détruiront en libérant les spermatozoïdes. Les spermatophores sont véhiculés le long d'un sillon vers l'extrémité du bras hectocotyle et déposés dans les oviductes* par des séries de mouvements péristaltiques du bras (mouvement dit « arch and pump » par les auteurs anglophones). Cette opération se reproduit de nombreuses fois, avec des délais plus ou moins brefs. L'accouplement peut durer d'à peine plus d'une minute à 3 heures, il est aussi extrêmement calme. Il peut être statique, ou se produire pendant que la femelle circule lentement à la recherche de proies. L'accouplement peut aussi se faire avec des partenaires en contact, le mâle étant au-dessus de la femelle et l'enveloppant de ses bras et de sa membrane brachiale, ce qui arrive notamment quand le mâle est plus puissant que la femelle et ne risque pas d'être tué par elle : deux cas de cannibalisme sexuel ont en effet été observés chez O. cyanea. Il est possible que le fait qu'O. cyanea, comme d'autres espèces, s'accouple à distance et à découvert soit lié à cette éventualité, la fuite du mâle en étant facilitée.
La ponte et la garde des œufs :
L'unique ponte des femelles a lieu un mois après l'accouplement. Elle s'étend sur plusieurs jours et se déroule dans un abri dont la femelle s'efforce de boucher l'entrée en utilisant les matériaux disponibles autour du nid. Les femelles pondent de 150 000 à plus de 700 000 œufs en plusieurs grappes reliées par un cordon et en une seule ponte. Les œufs sortant des oviductes pendant la ponte sont fécondés par les spermatozoïdes stockés. Ils sont oblongs et font environ 3 mm de diamètre. Les grappes, comprenant 600 à 1 200 œufs, sont fixées par une sécrétion adhésive sous le bloc qui sert d'abri ou sous la voûte d'une anfractuosité quelconque. La mère va ventiler ses œufs au moyen de son siphon respiratoire et les nettoyer avec le bout de ses bras durant toute la période d'incubation, qui dure entre 20 et 36 jours selon la température de l'eau. Si le poulpe est capturé et que les œufs ne sont plus nettoyés et ventilés, ils meurent rapidement par asphyxie du fait de leur recouvrement par les suspensions présentes dans l'eau. Ils peuvent aussi être la proie de murènes, de mérous, de crabes et même d'étoiles de mer, y compris en présence de la femelle.
La mort des géniteurs :
La femelle, qui a déjà cessé de se nourrir jusqu'à deux semaines avant la ponte, ne mangera plus pendant toute cette période et elle mourra généralement d'épuisement et de faim dans les 10 jours suivant la dernière éclosion. Cependant, contrairement à une idée reçue, cette mort peut survenir dans des délais très variables au sein d'une période de deux mois après l'éclosion des œufs, dans la mesure où il arrive que certaines d'entre elles recommencent à se nourrir à ce moment-là.
Les mâles entrent en sénescence après leur maturité sexuelle et leur période d'activité reproductrice. Il semble que ce sort soit lié à la glande optique, qui contrôle la reproduction et dont l'hyperactivité après la copulation déclencherait la sénescence. Ce déclin se caractérise par une perte progressive d'appétit, une hyperactivité désordonnée, des lésions blanches sur le corps, l'effet le plus spectaculaire étant une perte du contrôle des mouvements du corps. Cette perte affecte notamment les bras, qui partent en tous sens en se gênant mutuellement, et peut mener à l'autophagie, le poulpe dévorant ses tentacules. Les rares femelles qui survivent à l'éclosion de leurs œufs entrent de la même façon en sénescence. La durée de vie maximale de l'espèce se situe entre 12 et 15 mois.
Les juvéniles :
Le juvénile sort de son œuf en utilisant la propulsion créée par des contractions du manteau, et nage immédiatement en pleine eau pour rejoindre le large.
Il n'y a pas de stade larvaire*, le juvénile a l'aspect d'un petit poulpe dès l'éclosion. Cependant, ses bras sont beaucoup plus courts que le manteau, le rapport étant de 1 à 3, et il porte un entonnoir proportionnellement énorme en position ventrale. On parle alors de « paralarve ». Son corps est couvert de soies minuscules, il porte de petits points brun foncé et ses bras ne sont encore armés que de 3 à 4 ventouses par rangée. Il fait 3 mm de long et mène une vie planctonique* « paralarvaire » durant environ 30 à 40 jours, fonction de la température de l'eau, en se nourrissant de zooplancton* (notamment de larves de crustacés). La paralarve chasse à l'approche, elle peut déjà changer d'apparence en pâlissant ou s'assombrissant, et projeter de l'encre pour sa protection. A la fin de cette période, elle se laisse tomber sur le substrat et commence son existence benthique. Le cephalopodium du juvénile mesure alors environ 1 cm, et ses bras 2 cm. Puis les bras s'allongent plus vite que le manteau et le nombre de ventouses augmente. Les ocelles caractéristiques de l'espèce mettront encore plusieurs jours pour apparaître. Trois semaines après son installation*, le juvénile aurait une activité nocturne (vs l'activité diurne des adultes).
Peu de parasites internes, bactéries et virus ont été signalés ou étudiés chez O. cyanea. L'espèce est cependant parasitée par les copépodes exoparasites Octopicola stocki et Octopicola regalis.
Octopus cyanea peut collaborer avec un mérou dans un objectif de chasse : une étude de 2013 montre que le mérou Plectropomus leopardus signale la présence d'une proie inaccessible pour lui en se positionnant verticalement au-dessus de sa cachette et en faisant des mouvements saccadés de la tête, si un collaborateur potentiel se trouve dans un rayon de 10 mètres. Le collaborateur observé était un poulpe, O. cyanea. En 4 situations de signal sur 5, O. cyanea a répondu de façon appropriée au signal du mérou.
Variations de l'apparence : les possibilités de changement d'apparence chez O. cyanea, supérieures à celles de nombreux autres poulpes, sont d'autant plus développées qu'il est diurne. Ces variations intègrent des dimensions chromatiques (couleurs), texturales (imitation de la surface du support) et posturales (imitation du relief environnant), auxquelles peuvent s'ajouter des modifications de la posture de locomotion (imitation d'un poisson nageant). Elles forment un répertoire défini qui est caractéristique de l'espèce. Parmi leurs fonctions principales se trouvent la communication avec les conspécifiques, les stratégies de reproduction (attirer ou refuser un mâle, repousser ou berner un rival, etc.), la possibilité de se protéger de l'environnement (les radiations ultraviolettes, notamment), ainsi que la défense contre les prédateurs (camouflage ou intimidation) et l'approche des proies. Toutes les variations sont instantanées (la durée du changement est de l'ordre de la milliseconde) et peuvent se succéder à grande vitesse.
Sociabilité : O. cyanea est comme tous les poulpes un animal solitaire en dehors de la période de reproduction. La rencontre de deux individus peut tourner à l'affrontement. Celui-ci consiste d'abord en un entrelacement des bras à l'occasion duquel chacun se mesure par rapport à la puissance de l'autre, puis si nécessaire par un contact ventral pendant lequel chacun s'efforce de pousser l'autre, les morsures étant rares. Chez O. cyanea, on a observé que les vainqueurs prennent une livrée plus sombre que les vaincus. Le combat peut aussi devenir plus sérieux, les deux corps formant une boule compacte et la mort de l'adversaire semblant recherchée, souvent par asphyxie. Quand de nombreux individus se trouvent concentrés sur un même territoire de chasse, les plus grands ont un comportement dominant devant les plus petits, lesquels adoptent un comportement de subordonnés.
Défense contre les prédateurs : O. cyanea peut être la proie de murènes, de congres, de carangues, de requins, de dauphins, de tortues et de phoques. En général, quand les adultes sont attaqués ils s'en sortent avec un bras coupé, éventuellement volontairement, ce qui suffit souvent à contenter le prédateur. Quand il n'a plus le choix ou qu'il est surpris, il projette vers l'agresseur au moyen du siphon un ou plusieurs jets d'encre qui se diluent dans l'eau et provoquent un important nuage noirâtre faisant écran un certain temps (cela peut aller jusqu'à plusieurs minutes, fonction de l'hydrodynamisme) pour protéger sa fuite. Ce nuage, qui mêle encre et mucus, produit aussi des pseudo-formes susceptibles de tromper ou d'effrayer l'adversaire tout en altérant ses sens visuel et olfactif s'il attaque une pseudo-forme ou si le courant le met en contact avec l'encre. Le constituant principal de l'encre est une forme très concentrée de mélanine, un pigment dérivé d'un amino-acide, la tyrosine, à l'origine de l'altération des facultés sensorielles de l'agresseur. Quand l'animal est acculé, il couvre complètement sa tête et son manteau de ses bras pour former une sorte de boule, exposant à l'adversaire toutes ses ventouses et sa bouche cornée. Avant cela, il peut s'enfouir dans le sable en repoussant devant lui le sédiment déplacé par ses tentacules. Il arrive que le prédateur soit un autre poulpe, y compris un conspécifique, le cannibalisme étant référencé pour de nombreuses espèces de Céphalopodes.
Pupille horizontale : la pupille d'O. cyanea reste en position horizontale quelles que soient les positions du corps parce qu'elle est gérée par le système nerveux central à partir des informations données par les statocystes*, qui contrôlent notamment l'équilibre.
Hors d'eau : Les poulpes sont incapables de respirer à l'air, mais certaines espèces, dont O. cyanea, peuvent passer un certain temps hors de l'eau. Les chercheurs connaissent la tendance de certaines espèces à quitter leur aquarium et à traverser une pièce en rampant sur le sol pour rejoindre un autre bac contenant des poissons et les dévorer mais l'aventure est dangereuse : les fugitifs mourront au bout de 10 minutes si un bac n'est pas trouvé avant.
Sang bleu : son sang contient de l'hémocyanine (vs hémoglobine), une protéine riche en cuivre qui est dissoute dans le plasma et dont l'oxydation rend ce sang bleu (vs le fer oxydé de l'hémoglobine qui rend le sang rouge). La capacité limitée de l'hémocyanine à fixer l'oxygène est supposée être l'une des raisons de la lenteur relative du poulpe en dehors des comportements de fuite ou d'attaque.
Régénération : les bras coupés, volontairement (les poulpes sont capables d'autotomie*) ou pas, peuvent se régénérer. Les bras régénérés sont plus petits que leurs prédécesseurs, leur coloration est moins soutenue du fait d'un moins grand nombre de chromatophores, et les ventouses sont de moindre taille, quand elles ne sont pas difformes.
Ventouses : ses ventouses sont constituées d'une paroi musculaire ayant son propre système régulateur et sont indépendantes les unes des autres. Elles peuvent accomplir des actions stéréotypées sans ordre du système nerveux central. Leur constitution à deux chambres leur permet de créer un effet de succion rendant l'adhérence puissante et durable en générant des pressions négatives pouvant aller jusqu'à – 650 hectopascals. Ses ventouses rendent à O. cyanea de multiples services : capture des proies, fixation aux substrats durs, nettoyage du corps, locomotion, reconnaissance chémotactile des proies cachées ou des partenaires, manipulations d'objets, intimidation d'un adversaire ou séduction d'une partenaire, etc.
Autophagie : le poulpe est capable d'autotomie mais aussi d'autophagie, ce qui est très différent. Ce comportement n'est pas encore documenté chez O. cyanea, mais il est probable, dans la mesure où il est connu chez son supposé proche parent O. vulgaris. Ce processus d'autophagie peut avoir deux causes. La première est liée à la maturité sexuelle et à l'activité reproductrice via la sénescence qui lui succède (voir au paragraphe Reproduction), la seconde n'est pas clairement élucidée : il s'agirait de l'effet d'une substance pathogène produite par l'animal lui-même, ou plus probablement d'une maladie infectieuse attaquant le système nerveux. L'incubation prendrait entre une et deux semaines et la mort surviendrait un à deux jours après le commencement du comportement d'autophagie. Cette maladie peut surgir à n'importe quel âge.
Intelligence : O. cyanea est un animal remarquablement intelligent. Il a été démontré que les Céphalopodes sont capables d'exploration, de réactivité, d'accoutumance, de jeu exploratoire, de mémoire, de réactions émotionnelles, d'adaptation à des circonstances nouvelles, de prises de décision, d'apprentissage par association et observation incluant des aptitudes de discrimination visuelle et tactile. Ces compétences leur permettent de développer des comportements de résolution de problèmes pouvant aller jusqu'à l'usage d'outils.
« Conscience » : un certain nombre de chercheurs estime que les poulpes, parmi d'autres animaux, possèdent les « substrats neurologiques » (structures neuroanatomiques, neurochimiques et neurophysiologiques) requis pour des états de conscience « simples ». Au point que de nombreux spécialistes des neurosciences ont signé, en juillet 2012, un document intitulé la Déclaration de Cambridge sur la Conscience affirmant que l'homme n'est pas le seul animal à faire l'expérience d'états de conscience et d'états affectifs. Ce document mentionne les poulpes. http://fcmconference.org/img/CambridgeDeclarationOnConsciousness.pdf
« Personnalité » : on reconnaît à un certain nombre de poulpes bien étudiés des différences individuelles de comportement au niveau de l'activité, de la réactivité et de l'évitement, y compris en situations expérimentales standardisées, telles que l'on peut parler de « tempéraments » ou de « personnalités » (caractéristiques individuelles permanentes apparaissant tôt dans le développement). Son excellente mémoire à court et long terme permet de surcroît à O. cyanea de s'accoutumer à une présence non hostile. On l'a même vu rechercher ce contact en laboratoire : nombreux sont les chercheurs qui témoignent de comportements qu'ils assimilent à ceux d'un animal de compagnie.
Exigences éthiques de la recherche : l'ensemble de ces compétences a fait que les expérimentations sur des céphalopodes ont été placées en France et en Europe au même niveau d'exigence éthique que celles concernant les lapins, les souris, etc. Ainsi, la Directive 2010/63/UE du Parlement Européen et du Conseil du 22 septembre 2010 stipule que les céphalopodes devront être inclus dans son champ d'application au même titre que les vertébrés. Depuis le 1er janvier 2013, tous les projets de recherche impliquant des animaux vivants doivent de surcroît recevoir un avis favorable de la part d'un CEEA (Comité d'éthique en expérimentation animale). La Nouvelle-Zélande et certaines régions australiennes (le Territoire de la Capitale Australienne et l'Etat du Queensland) ont aussi étendu ce type d'exigences aux poulpes et aux calmars (ainsi qu'aux crustacés). De même, le Canada, la Suisse et la Norvège ont légiféré au sujet de l'utilisation des céphalopodes en laboratoire.
Pêche : les poulpes sont pêchés depuis toujours par les peuples côtiers, pour la richesse en protéines de leur manteau du point de vue de la subsistance, et pour la source de revenus qu'ils représentent du point de vue économique. Les pays asiatiques en pêchent et importent de très grandes quantités (le Japon notamment, pour la confection des sushis). Madagascar, entre autres, exporte O. cyanea. Le record de poids d'une prise d'O. cyanea à Hawaï en 2003 est de 8,623 kg.
L'UICN (Union Internationale pour la Conservation de la Nature) n'a pas encore défini le statut d'O. cyanea pour sa Liste Rouge. Sa maturité sexuelle précoce et sa grande fécondité font considérer l'espèce comme résistante, relativement à la pression de pêche. Cependant, dans un certain nombre de zones, des prélèvements irresponsables font craindre un fléchissement des populations d'O. cyanea (notamment du fait que la distinction entre mâles et femelles n'est pas faite, et que la surpêche des secondes peut tarir rapidement la ressource). De nombreux pays s'efforcent de contrôler la pêche au moyen de quotas, de périodes d'autorisation définies, de tailles minimales, d'interdiction de certaines techniques de pêche, etc. et en luttant contre la pêche illégale. Ces mesures sont en général inefficientes du fait du manque de moyens pour les faire appliquer, de l'étendue des zones à contrôler et de la pauvreté de populations dont cette pêche est souvent une importante source de subsistance.
Poulpe de récif commun : le mot poulpe vient du grec ancien [polypous], qui signifie « plusieurs pieds » (l'usage moderne parle de bras). La mention des récifs vient de ce que l'on trouve principalement cette espèce en milieu corallien, donc dans des biotopes récifaux. L'adjectif « commun » est justifié par les fortes densités observables de l'animal et par sa vaste distribution.
Octopus : du grec ancien [okto], qui signifie « huit », et [poús], qui signifie « pied ». Ces « huit pieds » renvoient évidemment aux huit bras des poulpes.
Le genre est décrit par Cuvier en 1797, il comprend actuellement 71 espèces dont les taxons sont acceptés. Il est caractérisé par deux rangées de ventouses sur chaque bras. Il est considéré comme un genre « fourre-tout » par beaucoup de taxonomistes, et devrait connaître des remaniements. O. cyanea, notamment, représente probablement un « complexe » d'espèces et pourrait même être placé dans le genre Abdopus. Le type* du genre Octopus est O. vulgaris.
cyanea : dans la mythologie grecque, Cyané [Kuanè] est une Naïade, la plus belle des nymphes aquatiques de Sicile. Le descripteur, J. E. Gray, a ainsi donné des noms issus de la mythologie grecque, y compris des noms de nymphes, à de nombreuses autres espèces, notamment de poulpes : Octopus aegina (la nymphe Egine), O. geryonea (Géryon), O. aranea (Arakhnê), O. eudora (la Néréïde Eudora), O. cassiopeia (Cassiopée), O. polyzenia (Polyxène), etc. Il est donc très probable que le nom d'espèce se réfère à la nymphe Cyané plutôt qu'à la couleur bleue.
On lit parfois que ce nom d'espèce, cyanea, vient du grec ancien [kuanos], qui désigne un minerai bleu foncé utilisé par les teinturiers, le terme moderne cyan désignant un bleu sombre. Cette explication semble peu probable : d'une part, cet animal ne présente que rarement quelques reflets bleutés dans sa livrée bordeaux, d'autre part et surtout, le descripteur a travaillé sur des spécimens conservés dans l'alcool qui avaient en conséquence perdu leur couleur. Celle-ci n'est d'ailleurs pas mentionnée dans sa description.
L'espèce est décrite par John Edward Gray (1800-1875), zoologiste au British Museum de 1824 à 1874, dans le Catalogue of the Mollusca in the collection of the British Museum, Part 1, Class IV, p. 15.
Le type vient d'Australie et est déposé au Natural History Museum de Londres.
Nous sommes redevables de la piste de recherche sur l'origine mythologique du nom d'espèce à Pauline Fiene (Mike Severns Diving, Hawaï).
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Cephalopoda | Céphalopodes | Yeux complexes, coquille interne, externe cloisonnée ou absente, cavité palléale musclée, siphon musculeux, tentacules ou bras (munis de ventouses). |
| Sous-classe | Coleoidea | Coléoïdes | Ventouses sur les bras, au nombre de 8 au moins. Bras hectocotyle chez le mâle pour le transfert des spermatophores. 2 branchies, 2 néphridies, une coquille interne ou vestigiale, des chromatophores, une poche à encre, un grand cerveau. |
| Super ordre | Octobrachia | Octopodiformes | Céphalopodes à huit bras, connectés par une sangle interbrachiale. Corps en sac globuleux. Ventouses rondes symétriques sans anneau corné. Jamais de crochets. |
| Ordre | Octopoda | Octopodes | 8 bras. Corps en forme de sac. Ce sont les poulpes. |
| Sous-ordre | Incirrina / Incirrata | Incirrates | Nageoires et lame interbrachiale perdues. Benthiques pour la plupart. |
| Famille | Octopodidae | Octopodidés | |
| Genre | Octopus | ||
| Espèce | cyanea |
 Céphalopodes
Céphalopodes

Un poulpe de taille moyenne
Octopus cyanea est un poulpe de taille moyenne. La livrée rouge foncé à bande médiane blanche est l’une des premières à apparaître quand l’animal s’estime dérangé.
Lagon de l'Ermitage, La Réunion, 1,50 m
19/03/2009
Céphalopodes

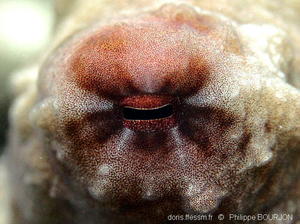
Ocelles
Sur cette livrée rouge ordinaire, on distingue à peine l’un des ocelles ovales situés entre les yeux et les paires de bras II et III. Il est usuellement beaucoup plus foncé et entouré d’un anneau clair bordé de brun. Ici, l’anneau est à peine plus clair que la livrée, et l’ocelle à peine plus foncé. Quoique caractéristiques de l’espèce, ces ocelles sont relativement rarement observés, l’animal les faisant disparaître à volonté.
Safaga, Egypte, mer Rouge, 12 m
06/10/2010


Livrée claire
La livrée claire peut aller jusqu'au blanc cassé ou au jaunâtre uniforme. Ce qui permet souvent, comme c'est le cas ici, de mieux voir les ocelles. Cet individu vient de changer de livrée : on voit encore des traces de la livrée rouge sur son corps.
La Réunion, 2 m
09/03/2011


Rapport bras-manteau
De nombreux poulpes sont difficiles à identifier in situ : le rapport entre la longueur du manteau et celle des bras peut y aider. Chez O. cyanea, les bras mesurent quatre fois la longeur du manteau
Lagon de l'Ermitage, La Réunion, 1,50 m
19/01/2011


Variation texturale hirsute
Cet individu ne doit pas être tout à fait d’accord pour poser (il est manifestement en train de chasser en mode « spéculatif »), ce pourquoi il adopte une livrée hirsute, les papilles étant hérissées : cette variation peut imiter les aspérités de l’environnement de façon à se rendre invisible, ou être destinée à décourager un adversaire.
Ce poulpe, en début de nuit, a visiblement été dérangé par les phares des plongeurs !
Sulawesi Nord, Indonésie, 12 m, début de nuit
14/04/2008


Bête à cornes
L’érection des papilles dans le but d’impressionner un adversaire peut aller jusqu’à doter soudain de « cornes » le poulpe stressé. Ces cornes peuvent se situer devant les yeux, comme sur cet individu, ou au-dessus. Elles se rétractent aussi vite qu’elles sont apparues si la tentative échoue.
Lagon de l'Ermitage, La Réunion, 1,50 m
20/04/2011

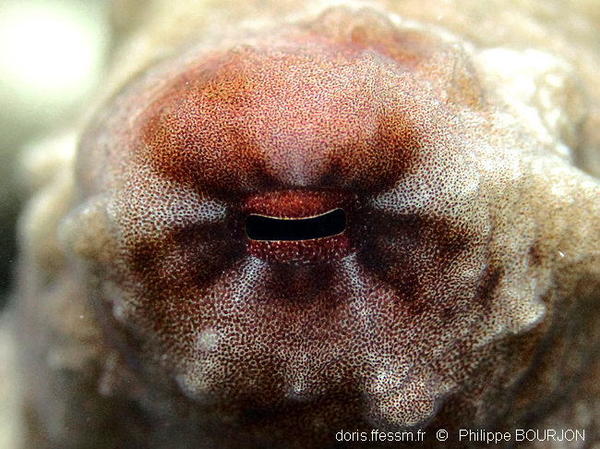
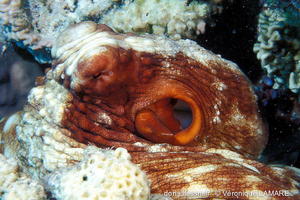
Œil
L’œil étrange des poulpes possède une pupille horizontale. Elle le reste quelle que soit la position de l’animal parce qu’elle est gérée par le système nerveux central à partir des informations données par les statocystes*, qui contrôlent notamment l’équilibre.
Lagon de l'Ermitage, La Réunion, 1,50 m
12/12/2012

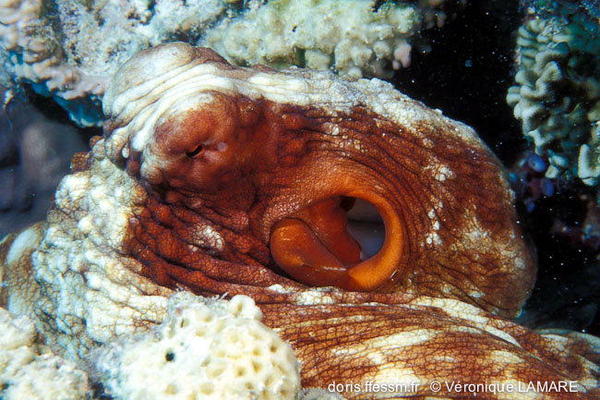
Cavité palléale
Ce poulpe, en train d'aspirer l'eau pour l'envoyer aux branchies, découvre ce faisant la cavité palléale et permet de distinguer le tissu musculaire enveloppant les viscères de l'animal.
Mer Rouge (Egypte)
16/07/2000


Respiration
L’eau est aspirée dans la cavité palléale, qui abrite les branchies, par un mouvement d’extension du manteau des deux côtés de la tête alors que l’entonnoir se referme. Elle est ensuite rejetée après échange gazeux dans les branchies via cet entonnoir, qui s’ouvre pendant que la cavité palléale se ferme des deux côtés grâce à un gonflement de la partie interne du siphon.
Lagon de l'Ermitage, La Réunion, 1,50 m
06/03/2011


Changements de livrée
Ces quatre photos représentent le même individu réagissant à l’approche de l’observateur :
Il est en train de chasser et porte d’abord la livrée unie bordeaux foncé typique d’Octopus cyanea quand il est calme, puis il pâlit en s’efforçant de prendre les tonalités de son environnement immédiat afin de s’y rendre invisible. Devant l’échec de sa tentative il arbore ensuite une bande blanche médiane susceptible de surprendre l’agresseur et enfin il montre des éléments de la livrée d’intimidation en train de se former, ce qui signifie qu’il est plus furieux qu’apeuré. On peut d’ailleurs constater qu’il reprend la partie de chasse troublée par le photographe.
Safaga, Egypte, mer Rouge
11/10/2009


La posture... mais pas la couleur
La posture inhabituelle de ce poulpe s’efforce d’imiter les reliefs environnants, mais il semble avoir oublié leurs couleurs…
Safaga, Egypte, mer Rouge, 20 m
1998


Intimidation
L’attitude d’intimidation la plus classique consiste à écarter les trois premières paires de bras (et ainsi à tendre la membrane brachiale) devant la tête et sur les côtés du manteau, chaque bras étant enroulé sur lui-même en spirale, tout en prenant une couleur rouge sombre à marques blanches.
Cet individu, que le photographe suivait depuis quelques minutes, a soudain fait face en prenant instantanément cette livrée, manifestant ainsi sa colère !
Lagon de l'Ermitage, La Réunion, 1,50 m
19/01/2011


Attaque parachute
Cette méthode de chasse est dite par les spécialistes chasse spéculative (du latin [speculare], épier, chercher) ou attaque parachute : le poulpe enveloppe de sa membrane brachiale déployée une zone susceptible de contenir des proies, qui s’y trouvent ainsi piégées. Puis les extrémités des bras fouillent la zone captive à la recherche d’organismes que l’animal n’a pas besoin de voir, puisque ses ventouses sont équipées de chémorécepteurs lui permettant de les identifier comme comestibles ou pas.
Lagon de l'Ermitage, La Réunion, 1,20 m
12/12/2012


Chasse spéculative
Il peut y avoir chasse spéculative sans attaque parachute : on observe fréquemment un poulpe posé sur le substrat ou à côté d’une roche en train d’explorer des anfractuosités sans que la membrane soit utilisée.
Wadi Lahami, sud Egypte, mer Rouge,
01/06/2009

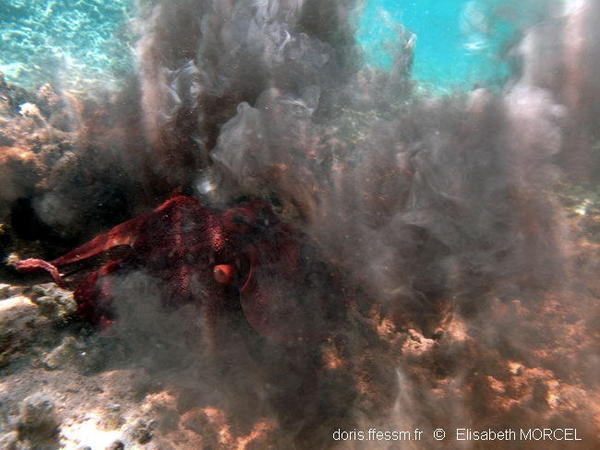
Jet d'encre !
La dernière solution du poulpe quand il s’estime en danger est d’envoyer un puissant jet d’encre mêlée de mucus pour faire un écran derrière lequel il fuit. Ce mélange est toxique pour les capacités visuelles et olfactives de l’agresseur attaquant une des « pseudo-formes » créées par le mucus, qui se disperse plus lentement que l’encre.
La cause du jet d’encre photographié n’a pas été vue par la photographe (le poulpe recule d’ailleurs dans sa direction).
Cette réaction étant coûteuse en énergie pour l’animal, il est préférable de ne pas tenter de la provoquer…
La Réunion, 2 m
18/11/2010


Locomotion
Comme les autres poulpes, Octopus cyanea peut « marcher ». Cette marche se fait généralement en se dressant, le manteau en position oblique à une trentaine de centimètres du fond, et en se servant de la courbe formée par les bras repliés sous son corps comme de « pieds », les ventouses étant orientées vers le substrat. L’allure est alors assez massive et pataude.
L’individu photographié ici montre que cette marche peut être plus élégante, le manteau étant plus haut, parallèle au substrat et seule l’extrémité des bras servant de pieds.
On a observé une autre espèce (Amphioctopus marginatus) transportant des demi-coques de noix de coco pour s’en faire un abri, marcher sur l’extrême pointe des bras comme sur des échasses (« stilt walking », écrit le chercheur australien).
Safaga, Egypte, mer Rouge, 12 m
27/05/2008


De nuit (sommeil)
Octopus cyanea est une des rares espèces de poulpes diurnes, bien qu'il se nourrisse préférentiellement au petit matin et au coucher du soleil. Il dort donc la nuit, et il le fait paupières closes, lové dans ses propres bras. Cet individu présente une tache blanche bordée de noir étonnante au-dessus du globe oculaire.
Lagon de l'Ermitage, La Réunion, 1,50 m
15/12/2010

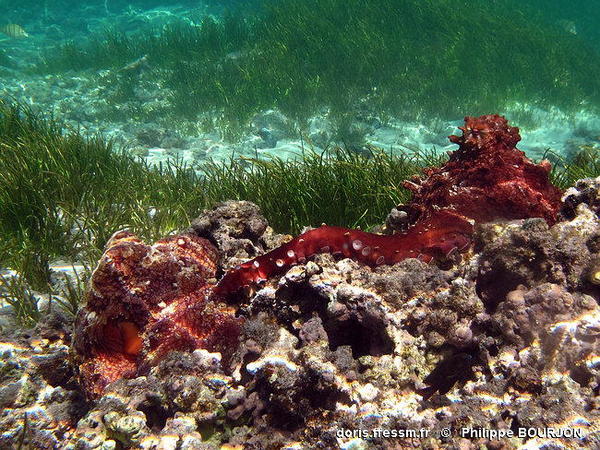
Accouplement
Lors de l'accouplement, le mâle (à droite, en livrée bordeaux à taches claires) introduit son tentacule reproducteur, l'ectocotyle*, dans la cavité palléale de la femelle (en livrée plus camouflée).
Lagon de l'Ermitage, La Réunion, 1,20 m
07/12/2008


Dépôt des spermatophores
L’extrémité du bras hectocotyle* a atteint son but : elle se trouve dans la cavité palléale de la femelle et dépose les spermatophores du mâle dans les oviductes de sa partenaire.
Beacon reef, îles Similan, Thaïlande, 27 m
11/04/2010


Grand juvénile
Le manteau de ce grand juvénile peut être estimé à 5 à 6 cm. Ses proportions sont à cette taille les mêmes que celles de l’adulte, ainsi que sa capacité de faire varier son apparence. On le voit ici développer la livrée d’intimidation sans prendre la posture qui lui est souvent associée (les bras enroulés sur eux-mêmes et entourant le manteau).
Lagon de l'Ermitage, La Réunion, 1,50 m
18/08/2013


A Mayotte
Cet individu photographié à Mayotte n’apprécie pas la proximité du photographe, comme le montre sa livrée.
Passe Bouni, Mayotte, 8 m
08/12/2008
Rédacteur principal : Philippe BOURJON
Vérificateur : Alain-Pierre SITTLER
Responsable régional : Alain-Pierre SITTLER
Alupay J. S., Caldwell R. L., 2013, The costs and benefits of losing an arm: autotomy in the octopus Abdopus aculeatus, Society fot Integrative and Comparative Biology, 2013 Annual Meeting.
Anderson R. C., Wood J. B., Byrne R. A., 2002, Octopus senescence : The Beginning of the End, Journal of Applied Animal Welfare Science, 5(4), 275-283.
Anderson R. C., Mather J. A., 2008, It's all in the cues: octopuses (Enteroctopus dolfleini) learn to open jars, Proceedings of the 3rd international Symposium Coleoid Cephalopods Through Time, Ferrantia, Travaux scientifiques du Musée d'Histoire Naturelle du Luxembourg, 8-13.
Boal J. G., 2006, Social recognition: a top down view of cephalopods behaviour, Vie et milieu – Life and Environment, 56(2), 59-79.
Bourjon P., Sittler A.-P., Noël P., 2016, Le poulpe de récif commun Octopus cyanea Gray, 1849, in Muséum national d'Histoire Naturelle [Ed.], Inventaire national du Patrimoine Naturel, pp. 1-24. Téléchargement
Budelmann B. U., 1998, Autophagy in Octopus, South African Journal of Marine Science, 20(1), 101-108.
Forsythe J. W., Hanlon R. T., 1997, Foraging and associated behavior of Octopus cyanea Gray, 1849 on a coral atoll, French Polynesia, Journal of Experimental Marine Biology and Ecology, 209(1-2), 15-31.
Grasso F. W., Setlur P., 2007, Inspiration, stimulation and design for smart robot manipulators from the sucker actuation mechanism of cephalopods, Bioinspiration and Biomimetics, 2, 170-181.
Hanlon R. T., Forsythe J. W., 2008, Sexual cannibalism by Octopus cyanea on a Pacific coral reef, Marine and Freshwater Behaviour and Physiology, 41(1), 19-28.
Hu M. Y., Yan H. Y., Chung W. S., Shiao J. C., Hwang P. P., 2009, Acoustically evoked potentials in two cephalopods inferred using the auditory brainstem response (ABR) approach, Comparative Biochemistry and Physiology, Part A, 153, 278-283.
Ibañez C., Keyl F., 2010, Cannibalism in Cephalopods, Reviews in Fish Biology and Fisheries, 20, 123-136.
Kier W. M., Smith A. M., 2002, The structure and adhesive mechanism of octopus suckers, Integrative and Comparative Biology, 42(6), 1146-1153.
Mather J. A., 2007, Cephalopods consciousness: behavioural evidence, Consciousness and Cognition, 17, 37-48.
Mather J. A., Anderson R. C., 1993, Personalities of Octopuses (Octopus rubescens), Journal of Comparative Psychology, 107(3), 336-340.
Sweeney M. J., Roper C.F.E., Mangold K. M., Clarke M. R., Boletzky S. W., 1992, “Larval" and juveniles cephalopods: a manuel for their identification, Smithsonian Contributions to Zoology, 513, 282p.
Van Heukelem W. H., 1976, Growth, bioenergetics and life-span of Octopus cyanea and Octropus maya, Dissertation de Doctorat, Université de Hawaï, 224p.
Villanueva R., Norman M. D., 2008, Biology of the planktonic stages of benthic octopuses, Oceanography and Marine Biology: An annual review, 45, 105-202.
Voight J. R., 1991, Enlarged suckers as an indicator of male maturity in Octopus, Bulletin of Marine Science, 49(1-2) 98-106.
Wood J. B., Anderson R. C., Interspecific evaluation of Octopus escape behavior, Journal of Applied Animal Welfare Science, 7(2), 95-106.
Yarnall J. L., 1969, Aspects of the behaviour ou Octopus cyanea Gray, Animal Behaviour, 17(4), 747-754.
Yoshida M., Ogura A., 2011, Genetic mechanisms involved in the evolution of the cephalopod camera eye revealed by transcriptomic and developmental studies, BMC Evolutionary Biology, 11, 180.
La page d'Octopus cyanea dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.




















")

