

Deux valves inéquilatérales pouvant mesurer plus de 50 cm de longueur
Valves fines avec 6 à 7 côtes radiales basses et larges à l’extrémité pointue
Seuls les jeunes individus ont des écailles en forme de pétale
Ouverture pour le passage du byssus réduite
Manteau gris-brunâtre sans marques colorées, avec de grands tubercules
Manteau ne dépassant pas le bord de la coquille
Absence d’ocelles
Siphon inhalant entouré de tentacules
Bénitier de tevoro (en fait bénitier tévoro, car ce n'est pas un nom de lieu).
Devil clam (GB), Teufelsmuschel (D)
Tridacna (Chametrachea) mbalavuana Ladd, 1934
Tridacna (Tridacna) mbalavuana Ladd, 1934
Tridacna tevoroa Lucas, Ledua & Braley, 1990
Indo-Pacifique
Zones DORIS : ● Indo-PacifiqueTridacna mbalavuana est une espèce rare, elle a une distribution limitée aux îles Fidji, aux îles Tonga, aux îles Loyauté, sur la côte est de la Grande Terre en Nouvelle-Calédonie et dans la frange extérieure de la grande barrière de corail en Australie.
Tridacna mbalavuana vit sur les récifs coralliens, plus profondément que les autres espèces (- 20m, - 30 m) et apparemment cette espèce n’est pas présente dans des eaux peu profondes.
La coquille de Tridacna mbalavuana peut atteindre 50 cm de longueur (voire 56 cm). Elle présente seulement de faibles sculptures. Les deux valves* sont inéquilatérales*, avec une extrémité pointue, elles sont fines et portent des taches colorées sur les côtés. Les valves portent 6 à 7 côtes radiales basses et larges, peu marquées, à la marge arrondie. Seuls les jeunes individus portent des écailles en forme de pétale. L'ouverture pour le passage du byssus* est réduite. Cette espèce est proche de T. derasa.
Le manteau* gris-brunâtre, sans marques colorées, porte de grands tubercules*. Il dépasse de très peu le bord de la coquille. Celle-ci peut s'ouvrir largement. Le manteau est dépourvu d'ocelles* (ou organes hyalins*). Le siphon* inhalant* est toujours entouré de tentacules* proéminents généralement blancs. Cette espèce possède peu d'iridophores* (voir la rubrique "informations complémentaires").
L'extérieur de la coquille est blanc-grisâtre avec souvent des taches rouges vers les sommets, souvent masquées par des algues calcaires et des éponges. L'intérieur de la coquille est blanc porcelaine.
L’identification des différentes espèces de bénitiers est délicate et difficile. Les motifs, les couleurs du manteau* varient considérablement d’un individu à l’autre pour la même espèce. Certaines combinaisons de couleurs et de motifs peuvent ressembler à celles d’une autre espèce.
En plongée, les individus vivants sont vus par dessus, la forme, le nombre de côtes et les marges de la coquille, sa symétrie, la présence d’écailles, les motifs et l’extension du manteau, la présence de tentacules* autour du siphon* inhalant*, la présence d'ocelles*, la profondeur et le fait que la coquille soit encastrée ou non dans le substrat* sont les seuls critères disponibles.
La coquille vide peut montrer la présence de l’ouverture pour le passage du byssus*.
12 espèces de bénitiers sont décrites actuellement. Toutes ces espèces ont une distribution Indopacifique et vivent à faible profondeur dans les récifs coralliens ou à proximité.
Certaines espèces sont plus répandues que d'autres:
Quelques espèces sont capables de s’encastrer dans le substrat: Tridacna maxima, T. crocea, T. noae et T. elongatissima.
Par ordre d'importance de présence :
Tridacna maxima (Röding, 1798). Cette espèce ressemble beaucoup à T. noae et T. elongatissima et un peu moins avec T. squamosina et T. crocea.
La coquille bivalve de forme triangulaire, allongée, est épaisse et lourde. Elle mesure en général 15 à 20 cm dans sa plus grande dimension (mais peut atteindre 42 cm). Chaque valve, inéquilatérale*, porte 4 à 6 (généralement 5) côtes radiales basses mais larges, sur lesquelles on observe de nombreuses écailles (ou scutelles*) grandes et ondulées disposées de façon concentrique (les plus anciennes sont souvent usées donc seules celles présentes sur la partie supérieure sont visibles chez les individus adultes).
Sur une coquille vide, la ligne de charnière représente environ un tiers de la longueur de la coquille. Le ligament* est externe. L’orifice pour le passage du byssus est de taille variable.
L'extérieur de la coquille est généralement blanc cassé à crème parfois nuancé de jaune, l'intérieur est blanc.
Chez l’animal vivant, les bords du manteau recouvrent en général les bords de la coquille. Le siphon est bordé de petits tentacules. Les siphons et le manteau sont richement tachetés, pigmentés souvent de couleur vive (bleu, vert, brun) mais pas iridescents avec à peine deux motifs identiques. Le long de la marge du manteau, des ocelles sont disposés en rangées plus ou moins régulières et forment une ligne sombre presque continue sur le bord du manteau. Ils sont également dispersés de façon irrégulière sur le manteau. T. maxima n’est jamais aussi profondément encastrée que T. crocea.
C’est l’espèce de bénitier la plus commune, elle a une très large distribution. T. maxima est présente du nord de la mer Rouge (golfe de Suez et golfe d’Eilat/Aqaba), en englobant les récifs des océans Indien (dont les îles Éparses, Mayotte, La Réunion) jusqu'au Pacifique (dont la Nouvelle-Calédonie, Wallis-et-Futuna et la Polynésie française).
Tridacna squamosa Lamarck, 1819, Cette espèce est très commune (juste après T. maxima). La coquille peut mesurer jusqu’à 40 cm de longueur. Chaque valve est faiblement inéquilatérale*, et contrairement à T. maxima, elle porte 4, 5 ou 6 grandes côtes radiales arrondies espacées. De nombreuses grandes et épaisses écailles espacées sont présentes sur les côtes. Les bords de la coquille sont fortement incurvés. L’ouverture pour le passage du byssus est très variable (étroite à modérée).
Les valves sont souvent colorées (jaune et rose-orange).
L’extension du manteau peut cacher la coquille et les écailles. Le siphon inhalant est bordé de tentacules nombreux et complexes. Le manteau de T. squamosa présente généralement des motifs tachetés dans des combinaisons de jaune, orange, bleu, vert et brun. Le manteau est lisse avec parfois quelquese papilles.
Cette espèce est observée sur les platiers récifaux peu profonds jusqu'à une profondeur d'environ 20 m, voire jusqu’à 42 m dans les sites abrités. Elle peut être partiellement recouverte par la croissance du corail.
Elle est présente en mer Rouge, en Afrique de l'est, à Madagascar et aux Mascareignes* et Chagos, jusqu'aux îles Tonga, Tuamotu et l’archipel des Gambier et du sud du Japon au Queensland (Australie), sauf aux îles Cocos (Keeling) aux îles Mariannes du Nord où l'espèce est localement éteinte. Cette espèce a été introduite à Hawaï et à Guam.
Tridacna gigas (Linnaeus, 1758). C’est le plus grand mollusque bivalve vivant. Le plus grand spécimen mesurait 1,37 m de longueur et les 2 valves pesaient 230 kg. Les jeunes individus sont difficiles à distinguer des autres espèces de tridacnides, ils sont souvent confondus avec T. derasa.. La coquille est épaisse et lourde, sans écailles. Chaque valve est légèrement inéquilatérale par rapport à son sommet, elle porte 4 à 5 côtes radiales. L’ouverture pour le passage du byssus est petite ou absente.
L’extension du manteau est variable, il peut rester juste sur le bord ou cacher complètement la coquille. Le manteau est généralement brun doré, jaune ou vert avec de nombreuses taches iridescentes bleues, violettes ou vertes, surtout sur les bords. Les plus grands individus peuvent avoir tellement de taches que le manteau semble bleu ou violet uni. Le siphon inhalant ne porte pas de tentacules.
La coquille ne peut pas se fermer complètement.
T. gigas vit à faible profondeur voire dans des zones abritées de la zone de balancement des marées, et plutôt sur des récifs isolés en raison de sa surexploitation par l’homme. T. gigas, en Nouvelle-Calédonie, est présente à l'état de fossile, et est apparemment éteinte. T. gigas est présent de la mer Rouge au Pacifique central en passant par l'Afrique de l'Est et les îles Éparses, la Nouvelle-Calédonie et la Polynésie française.
Tridacna derasa (Röding, 1798). C’est la deuxième plus grande espèce puisque sa coquille peut atteindre jusqu’à 60 cm de longueur. Mais les coquilles sont plus allongées et étroites que celles de T. gigas. Les valves sont symétriques par rapport à leur sommet, et portent 5 à 7 côtes radiales peu développées. En général chaque valve porte peu d’écailles voire de simples crêtes. L’ouverture pour la passage du byssus est étroite et petite.
L’extension du manteau est variable, à peine au-delà de la marge pour cacher la coquille. Le siphon inhalant est bordé de grands tentacules divisés. Cette espèce perd son byssus et peut donc être libre.
Elle peut être observée jusqu’à 20 m de profondeur.
Tridacna mbalavuana ressemble à T. derasa, mais se distingue par la présence de tubercules* à la surface de son manteau et de tentacules autour de l’orifice du siphon inhalant, une coquille plus mince et des taches colorées sur les côtes.
L'aire de répartition de Tridacna derasa est centrée sur le triangle de corail, elle est également présente à l'ouest de cette zone jusqu'aux îles Cocos (mais pas dans l'Océan Indien à l'ouest des Cocos), en Nouvelle-Calédonie, au nord de cette zone jusqu'au sud du japon (archipel des Ryükyü) et à l'est jusqu'aux îles Tonga.
Tridacna noae (Röding, 1798), (= T. ningaloo). La coquille mesure entre 6 et 20 cm (voire jusqu’à 28 cm) de longueur. Les valves sont inéquilatérales avec 5 à 7 côtes radiales. Les écailles sont bien espacées.
Cette espèce ressemble à T. maxima en apparence, mais se distingue par ses ocelles peu distribués et espacés sur le bord du manteau et plus particulièrement par son ornementation de taches ovales (ou en forme de goutte d’eau) de différentes couleurs délimitées par des marges blanches le long du bord du manteau et par la présence de papilles sur le manteau.
Elle est observée de 1 à 15 m de profondeur souvent encastrée dans les substrats récifaux qu'elle serait capable de creuser.
Longtemps confondue avec T. maxima mais généralement présente en plus faible abondance, sa distribution confirmée est parcellaire, elle doit être revue. Tridacna noae est documentée dans l'océan Indien en Afrique de l'est (Tanzanie) et dans l'ouest Pacifique au Japon (archipel des Ryükyü) et à Taïwan, ainsi qu'aux Vanuatu.
Tridacna crocea Lamarck, 1819. C’est l’espèce la plus petite des bénitiers, elle est morphologiquement très proche de T. maxima et de T. squamosa mais la longueur de la coquille est de 10 cm (rarement jusqu’à 15 cm). Chaque valve est modérément inéquilatérale et modérément allongée. Elle porte 5 à 6 (parfois jusqu'à 10 avec 4 ou 5 côtes moins prononcées) côtes radiales basses avec de nombreuses écailles fines, petites très rapprochées les unes des autres, mais souvent érodées sauf celles près de la marge. L’ouverture byssale* est grande (1/3 de la longueur de la coquille).
L’extension du manteau peut cacher la coquille et les écailles. Le manteau est très richement pigmenté (habituellement bleu ou vert iridescent avec des mélanges de nombreuses autres couleurs comme du violet, de l'orange). Il présente des motifs variés, des rayures, des vagues, des taches et des anneaux. Le siphon inhalant est bordé de nombreux petits tentacules simples.
La coquille est blanc-grisâtre parfois teintée de jaune, d'orange. parfois une bande colorée est présente sur le bord supérieur de la coquille et sur sa surface intérieure.
Cette espèce peut être complètement encastrée dans le corail. Seuls les bords des valves peuvent affleurer la surface.
Elle semble supporter des eaux saumâtres et même l'émersion car elle vit dans la zone de balancement des marées et à faible profondeur (10 m maximum). De fortes densités ont été observées : 100 à 200 individus par m².
Elle est présente en Afrique de l'est, à Madagascar, dans les Mascareignes* et dans le triangle de corail, du Japon à l’Australie et des îles Andaman (océan Indien) jusqu'au Vanuatu.
Hippopus hippopus (Linnaeus, 1758). Cette espèce mesure jusqu’à 40 cm de longueur (voire 50 cm). Les valves épaisses, étroitement imbriquées, sont inéquilatérales (parfois une valve est plus creuse que l'autre). Elles portent 7 à 8 côtes radiales sans écailles avec des taches brunâtres ou rougeâtres. L’orifice pour le passage du byssus est étroit et disparaît chez l’adulte.
Le manteau brunâtre ou vert-olive avec des motifs marbrés ne s’étend pas au-delà des bords de la coquille. Il ne porte pas d’ocelles et le siphon inhalant ne possède pas de tentacules. Cette espèce perd rapidement son byssus et de ce fait peut être trouvée sur des fonds sableux ou d’herbiers peu profonds (jusqu’à 10 m de profondeur). Elle peut se déplacer en roulant par des contractions du muscle adducteur*.
Cette espèce est présente du sud du Japon au nord-est de l’Australie et de l’Indonésie aux îles Tonga.
Tridacna elongatissima Bianconi, 1856. La coquille, allongée, lourde, presque triangulaire, peut mesurer jusqu’à 34 cm de longueur. Les valves inéquilatérales portent 6 à 7 côtes radiales avec des écailles circulaires assez grandes et rapprochées plutôt limitées aux sommets. L’orifice pour le passage du byssus est large. L’extérieur de la coquille est blanc-grisâtre ou jaune et l’intérieur blanc porcelané*.
Le manteau est vert à brun.
Cette espèce est présente jusqu’à 12 m de profondeur seulement dans l’ouest de l’océan Indien (île de La Réunion, mais rare à l’île Maurice et apparemment absente à l’île Rodrigues et à Mayotte). Elle a été par le passé souvent confondue avec T. maxima avec laquelle elle vit.
Tridacna squamosina Sturany, 1899, (= T. costata, T. elonga). Cette espèce ressemble à T. maxima et T. squamosa. La coquille peut mesurer jusqu’à 32 cm de longueur. Les valves sont inéquilatérales avec 5 à 7 côtes radiales pointues à leur extrémité. Les écailles sont bien espacées près des sommets et serrées sur les bords. Les deux valves fermées laissent un espace. Le byssus est développé, l'ouverture pour son passage est moyenne.
Le manteau est très variable avec une large gamme de couleurs et de motifs et est couvert de petites papilles en forme de boutons. Le siphon inhalant est bordé par des tentacules
Tridacna squamosina vit vers 5 m de profondeur dans les zones récifales et les herbiers, elle est endémique* de la mer Rouge.
Cette espèce a été surpêchée lors des premières occupations humaines de la région (grand nombre de coquilles subfossiles -125 000 ans).
Tridacna rosewateri Sirenko & Scarlato, 1991 (= T. lorenzi). La coquille de 6 à 19 cm de longueur (jusqu’à 26 cm) est plus fine que celle de T. squamosa, et de T. maxima avec des prolongements des côtes plus développées, les valves sont faiblement inéquilatérales et elles portent 4 à 5 côtes radiales arrondies. Les écailles sont plus grandes et éparses. L’orifice pour le passage du byssus est assez large. L’intérieur des valves est blanc porcelané.
Cette espèce est endémique du plateau des Mascareignes* où elle a été observée parmi des coraux clairsemés dans les herbiers à Thalassodendron ciliatum au nord-est de l’île Maurice et dans l'archipel de St Brandon, qui appartient à la République de l'île Maurice.
Hippopus porcellanus Rosewater, 1982. Cette espèce, rare, possède une coquille plus lisse et plus fine que celle de H. hippopus. Elle mesure souvent moins de 40 cm de longueur. L’orifice pour le passage du byssus est étroit. Elle peut être confondue avec T. derasa mais le manteau est généralement gris ou brun, dépourvu d’ocelles, il ne s’étend pas au-delà des bords de la coquille. Le siphon inhalant possède des grands tentacules.
Cette espèce vit dans la zone de balancement des marées et peu profondément le long des bords des lagons. Sa distribution est restreinte aux Philippines, à l’Indonésie et à la Papouasie-Nouvelle-Guinée et dans le triangle de corail.
Au cours des 15 dernières années, trois espèces ont été redéfinies grâce aux outils moléculaires : T. squamosina, T. noae et T. elongatissima, toutes les trois morphologiquement proches d'autres espèces avec lesquelles elles avaient été mises en synonymes: dans ce cas on parle d’espèce cryptique*. Ce phénomène est assez commun chez les invertébrés marins et il existe peut-être encore d’autres espèces non décrites ou cryptiques"
Nicaisolopha tridacnaeformis (L.R. Fox, 1927). C’est une huître (famille des Ostreidés) dont les valves ressemblent un peu à celles des bénitiers. Cette espèce vit dans l'océan Pacifique
Les bénitiers sont proches des Cardiidés (comme les coques communes par exemple). Ces dernières sont des filtreurs*. Cependant les bénitiers comme T. mbalavuana ont une alimentation particulière. Le manteau* de ces mollusques contient de très grandes quantités de petites algues unicellulaires, les zooxanthelles* (Famille des Symbiodiniacées). Celles-ci sont des Dinoflagellés symbiotiques*. Les bénitiers, comme les coraux des récifs coralliens, se nourrissent des produits de la photosynthèse* de ces algues et de leur digestion. C’est la présence de ces algues symbiotiques qui est à l’origine de la distribution des bénitiers dans des eaux claires et à faible profondeur afin de permettre la photosynthèse. Ces zooxanthelles sont logées dans une structure ramifiée (appelée système tubulaire à zooxanthelles) qui communique avec l’estomac de l’animal.
Les bénitiers peuvent s'associer à plusieurs genres de Symbiodiniaceae, qui varient selon la distribution géographique et l’espèce.
Notamment,
seule T. mbalavuana, qui est l'espèce de bénitier la plus profonde, a
augmenté son efficacité photosynthétique dans les conditions d'éclairage les
plus faibles.
L'espèce
présente plusieurs adaptations morphologiques uniques qui augmentent
probablement son efficacité photosynthétique à des intensités lumineuses
réduites. Ces adaptations comprennent un manteau rugueux
et une ouverture inhabituellement large des valves, qui augmentent tous deux la
surface disponible pour que les zooxanthelles absorbent la lumière disponible.
En outre, la rareté des iridophores (c'est-à-dire des cellules à haute
réflectance communes dans le manteau des bénitiers) et la distribution
relativement peu profonde des zooxanthelles dans le manteau maximisent
probablement la disponibilité de la lumière pour les symbiotes*. Cependant,
malgré l'efficacité accrue de sa machinerie photosynthétique, des auteurs Klumpp et Lucas (1994) estiment qu'à la profondeur maximale de sa distribution,
la combinaison de l'alimentation en suspension et de la photosynthèse ne
fournit que 83% de la nutrition nécessaire pour maintenir la croissance de T.
mbalavuana. Les auteurs supposent donc que l'espèce doit avoir la capacité
d'accéder à d'autres sources de nutrition, telles que la matière organique
dissoute, pour combler la différence.
D’autres organismes exploitent les produits de la photosynthèse d’algues symbiotes* : une coque tropicale Corculum cardissa (bucarde cœur de vénus), des éolidiens comme la limace à bigoudis (Spurilla neapolitana), des sacoglosses comme certaines élysies (Elysia viridis), des gorgones, des hydraires, des méduses (comme Cassiopea andromeda par exemple) et bien entendu les madréporaires des récifs coralliens.
Les bénitiers sont des hermaphrodites* protandres*. La partie mâle de la gonade* se développe en premier, puis la partie femelle. Les cellules sexuelles (spermatozoïdes* et ovules*) maturent presque simultanément et sont libérées dans l’eau. La synchronisation de la libération de ces cellules est assurée par l’émission d’une substance induisant cette libération (une phéromone*). Elle est détectée par des chimiorécepteurs du siphon* inhalant*. L’animal va alors se gonfler d’eau, fermer son siphon inhalant puis en contractant son muscle adducteur* qui va fermer la coquille, il va expulser l’eau et les cellules sexuelles par le siphon exhalant*. Les spermatozoïdes sont émis en premier, ensuite les ovules (environ 100 µm de diamètre). L’autofécondation n’est pas possible. La fécondation a lieu en pleine eau.
Chaque œuf se développe, en environ 12 heures, en une larve* trochophore*. Celle-ci commence à sécréter une coquille de carbonate de calcium. Après deux jours, la larve* mesure 160 µm et devient une larve véligère*. Elle développe ensuite un pied. Après dix jours de vie planctonique*, la larve rejoint le fond et peut se déplacer avec son pied puis elle se métamorphose* en un bénitier juvénile.
Les larves de bénitiers ont tendance à s'installer sur des substrats* qui offrent un abri sous forme de rainures et de crevasses, Les bénitiers juvéniles finissent par s'attacher au substrat à l'aide des filaments du byssus*.
Le jeune tridacne ne possède pas encore de zooxanthelles*, il se nourrit de plancton*. Ce n’est qu’après une vingtaine de jours qu’il capture les zooxanthelles présentes dans la colonne d'eau..
La taille et l'âge
de la maturité varient selon l'espèce et l'emplacement géographique, mais en
général, les bénitiers sont connus pour atteindre la phase de maturité mâle
dès 2-3 ans et la phase de
maturité femelle vers 3-5 ans.
La durée de vie des petites et moyennes espèces de bénitiers serait de 10 à 50 ans.
En plus des zooxanthelles* contenues dans leurs tissus, les Tridacnidés servent d'hôtes à un grand nombre d'organismes qui vivent sur la surface des valves (épibiontes*), s'enfouissent dans l’épaisseur de la coquille ou encore habitent la cavité du manteau* (commensaux*, ectoparasites* et endoparasites*).
Parmi les épibiontes, certaines algues encroûtantes ou non, et également des éponges (comme les cliones), des anémones, des ascidies, des bryozoaires et des vers tubulaires peuvent parfois encrasser la coquille et nuire à la croissance du bénitier.
Certains même forent la coquille comme certaines éponges, des mollusques bivalves (Lithophaga et Gastrochaena), des annélides polychètes tubicoles* (Oenone fulgida), etc...
D’autres organismes commensaux vivent dans les cavités du manteau* comme plusieurs espèces de crevettes Pontoniinidées, des crabes Pinnothéridés, voire des petits poissons de récifs (comme Encheliophis homei) et même des poissons des anémones si celles-ci font défaut.
Différents parasites* sont également présents comme des copépodes* cyclopoïdes (Anthessius, Lichomolgus), des gastéropodes Pyramidellidés (ces derniers insèrent leur proboscis* dans le
tissu du bénitier et comsomment l’hémolymphe*).
Pour en savoir plus vous pouvez consulter l’article de Neo et al. 2015.
Les infections dues à des bactéries sont le plus souvent causées par des Rickettsia sp. qui infectent les branchies*. Celles causées par des protozoaires* sont souvent dues à des Marteila sp. ou Perkinsus sp.. Ce sont surtout les jeunes individus qui sont touchés.
Certains auteurs considèrent les Tridacninés comme des espèces ingénieures* car elles jouent de nombreux rôles écologiques sur les récifs coralliens (Neo et al. 2015.)
Chez un bivalve typique comme la coque commune (proche des bénitiers), les siphons* inhalant* et exhalant* sont situés à l’arrière de l’animal, les sommets de la coquille sont en haut et le pied en bas. Ce pied peut sortir entre les bords libres des deux valves de la coquille.
Un bénitier présente une organisation originale. Comme il doit permettre à la lumière d’éclairer les zooxanthelles*, il faut que les tissus contenant ces algues puissent être exposés à la lumière solaire. La charnière de la coquille est déplacée vers le bas près de l’ouverture par laquelle sortent les fils du byssus* (l’animal a subi une rotation de 180 ° dans sa coquille).
Les tissus contenant les zooxanthelles sont largement hypertrophiés et s’étendent sur toute la longueur de la surface supérieure couvrant l’ouverture entre les bords des deux valves afin d’offrir une grande surface d’exposition à la lumière solaire. A cause de ces modifications l’un des deux muscles adducteurs* a disparu (l’adducteur antérieur). Ainsi les bénitiers sont des monomyaires* (un seul muscle adducteur) comme les huîtres et les coquilles Saint-Jacques mais pour des raisons différentes.
Ces profondes modifications de l’anatomie des bénitiers se sont effectuées au cours des temps géologiques.
Ainsi la partie dorsale habituelle de la coquille est devenue partie ventrale et vice versa de telle sorte que l‘«orientation» d’une valve de bénitier est très particulière.
Comment déterminer chez les bénitiers les valves droites et gauches ? Il faut tenir un individu valves accolées l’une contre l’autre en plaçant le sommet et l’échancrure où passe le byssus en position inférieure et le sommet du côté opposé à l’observateur. La coquille ainsi disposée, on a face à soi les bordures ondulantes des deux valves qui représentent la bordure dorsale, la valve qui est à droite est la valve droite, celle qui est à gauche est la valve gauche (Salvat, Rives 1975).
Tridacna mbalavuana présente des caractères semblables à ceux du genre Hippopus comme l’absence d’une fente pour la passage du byssus, le manteau qui ne déborde pas dur les bords des valves et l’absence d’ocelles*.
Les Tridacnidae semblent s'être séparés des autres Cardiacés vers le début du Cénozoïque (l’ère tertiaire) occupant peut-être la niche laissée vacante par l'extinction, il y a 66 millions d'années, des bivalves rudistes (comme les Hippurites). Le genre Tridacna est apparu au cours de l’Yprésien (soit il y a 56 à 47 Millions d'années).
La sous-famille des Tridacnes (les Tridacninés) comprend deux genres :
La lumière du soleil est bien sûr essentielle à l'activité photosynthétique* des zooxanthelles* abritées dans le manteau*, mais elle est potentiellement dangereuse pour le bénitier. Le risque d'exposition à des longueurs d'onde nocives a été surmonté par la production d'une pigmentation protectrice. Ces pigments sont contenus dans des cellules appelées iridophores* dans les tissus exposés. Ces cellules pigmentaires contiennent des empilements de minuscules plaquettes qui ont plusieurs fonctions. Ici elles réfléchissent la lumière et ainsi génèrent des couleurs par diffraction de la lumière, c’est le même effet qui est observé sur la coquille d’autres mollusques, les plumes de certains oiseaux et les ailes de certains papillons. Ces iridophores sont donc à l’origine de la gamme de couleurs allant du bleu au vert ou du brun au jaune, ils donnent lieu à une variété presque infinie de motifs qui sont les plus vifs chez les plus petites espèces.
De nombreux prédateurs peuvent s’attaquer aux bénitiers.
Les juvéniles sont attaqués par des crabes et certains poissons comme le Pseudobalistes flavimarginatus qui peuvent écraser les valves de la coquille, ou des gastéropodes perceurs (Fasciolaridés, Muricidés) et des astéries.
Les marques de dents sur les surfaces extérieures de la coquille indiquent des attaques par des poissons de récif comme les poissons perroquets et des entailles sur les marges du manteau par les poissons chirurgiens.
Pour les individus plus âgés, les prédateurs potentiels comprennent des raies aigles Myliobatidés comme Aetobatis narnari, les tortues Caretta caretta, Chelonia mydas et certains grands poissons, mais leur impact diminue à mesure que les bénitiers grandissent (Adams et al. 1988).
Harry S. Ladd, en 1934, a fait la première description de Tridacna mbalavuana à partir de coquilles fossiles du Néogène (tertiaire supérieur soit entre 23 et 2 millions d’années) de l’ïle Vitilevu aux Fidji.
Aux îles Tonga, T. mbalavuanaa été récolté pour les marchés intérieurs (par plongée sous-marine ou méthodes de pêche traditionnelles).
Le réchauffement climatique a également des effets indésirables sur les populations de bénitiers.
Les bénitiers sont confrontés à un certain nombre de menaces naturelles et d’origine humaine dans l’ensemble de leur aire de répartition comme :Selon l'UICN*: espèce vulnérable
(B1+2c) soit : en danger d’extinction dans l’ensemble de son domaine de répartition.
Bénitier : depuis la Renaissance, le recours au tridacne comme bénitier est devenu traditionnel pour contenir de l'eau bénite à l'entrée des églises catholiques et de ce fait a donné au « tridacne », son nom usuel de « bénitier ». Toutefois, ce sont surtout les valves de Tridacna gigas qui sont utilisées.
« Bénitier » vient de l’ancien français benoitier puis benestier issu de eau benoitier lui-même dérivé de l’ancien français ewe benëeite, benëoite.
du diable : traduction du nom fidjien.
Tridacna : le zoologiste et médecin français Jean Guillaume Bruguière (1750-1798) ne donne pas d'explication. Ce nom de genre latin vient du grec [tridakna], [tri] = trois et [dakn] = mordre. Cailleux et Komorn (1981) traduisent ce mot par "qu'il faut mordre trois fois ". On trouve également "dont on fait trois bouchées". Tridacna a été utilisé par le scientifique romain Pline l'ancien (23-79) pour des coquillages de très grande taille (comme des huîtres par exemple). Un des noms espagnols de la grande nacre est tridacna.
mbalavuana : Ladd, en 1934, n’a pas donné d’explication. La règle d’antériorité des noms scientifiques a remplacé le nom Tridacna tevoroa, donné par Lucas, Ledua & Bradley en 1990, après sa redécouverte, par T. mbalavuana.
tevoroa : en fidjien [tevoro] = diable, tevoroa est la latinisation du nom fidjien.
Numéro d'entrée WoRMS : 466759
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Bivalvia / Lamellibranchia / Pelecypoda | Bivalves / Lamellibranches / Pélécypodes | Mollusques aquatiques, filtreurs, au corps comprimé latéralement. Coquille composée de 2 valves articulées disposées de part et d’autre du plan de symétrie. Absence de tête, de pharynx, de radula et de glande salivaire. |
| Sous-classe | Autobranchia | Autobranches | |
| Infra-classe | Heteroconchia | Hétéroconchie | |
| Subter-classe | Euheterodonta | Euhétérodonte | |
| Super ordre | Imparidentia | Imparidenties | |
| Ordre | Cardiida | Cardiides | |
| Super-famille | Cardioidea | ||
| Famille | Cardiidae | Cardiidés | Coquille fermée en forme de cœur. Côtes radiales fortes avec parfois présence de longs tubercules. Bord des valves crénelé. |
| Sous-famille | Tridacninae | Tridacninés | |
| Genre | Tridacna | ||
| Espèce | mbalavuana |


Un bel individu
La coquille largement ouverte, la couleur du manteau et ses gros tubercules, l'absence d'ocelles sont caractéristiques. On distingue à gauche le siphon exhalant et à droite bordé de tentacules blancs le siphon inhalant.
Touho, Côte est de la Grande Terre, Nouvelle-Calédonie, 30 m.
10/08/2015

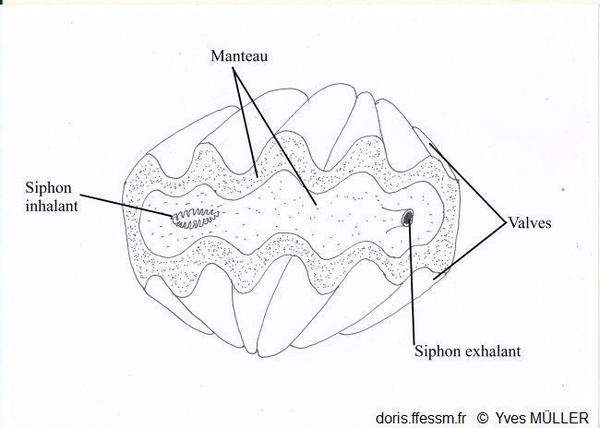
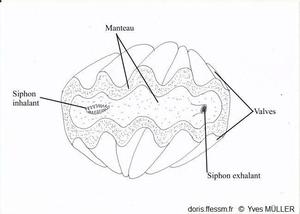
Un bénitier vivant vu de dessus
Le manteau et les siphons vus par-dessus.
Dessin
03/02/2023

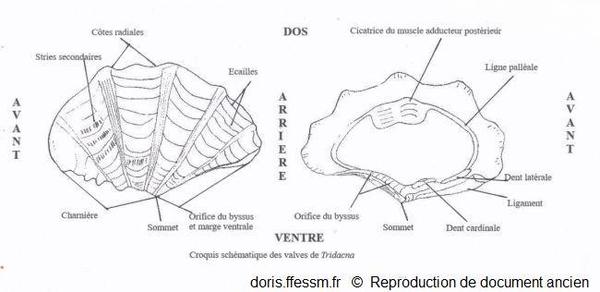
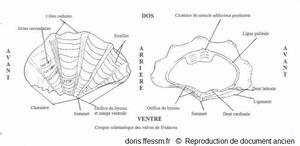
Schéma des valves de Tridacna
Ce schéma présente quelques caractéristiques des coquilles des bénitiers.
Schéma modifié d'après la planche 266 page 350 de Rosewater 1965
Reproduction de documents anciens
1965

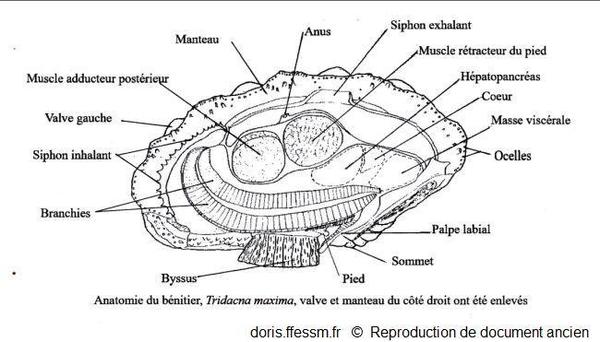
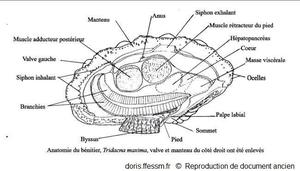
L'anatomie de tridacna maxima est identique à celle de T. mbalavuana
La valve et le manteau du côté droit ont été enlevés.
Schéma modifié d'après la planche 269 (page 352) de Rosewater, 1965
Reproduction de documents anciens
1965

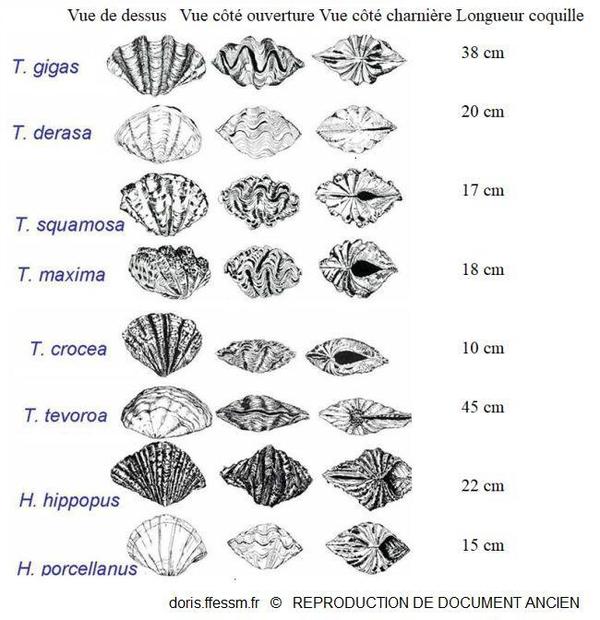
Quelques espèces de bénitiers
T. tevoroa = T. mbalavuana.
Sur cette planche, T. noae, T. elongatissima, T. squamosina et T. rosewateri ne sont pas représentés.
D’après les planches 13 (page 19) et 14 (page 20) de Norton et Jones, 1992
Reproduction de documents anciens
1992

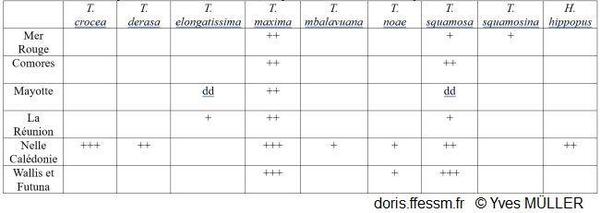
Distribution des bénitiers dans les domaines géographiques utilisés dans DORIS
+++ = abondant, ++ = fréquent, + = rare, dd = données déficientes.
D’après Neo & al 2017 et Fauvelot C (com. pers).
21/11/2023

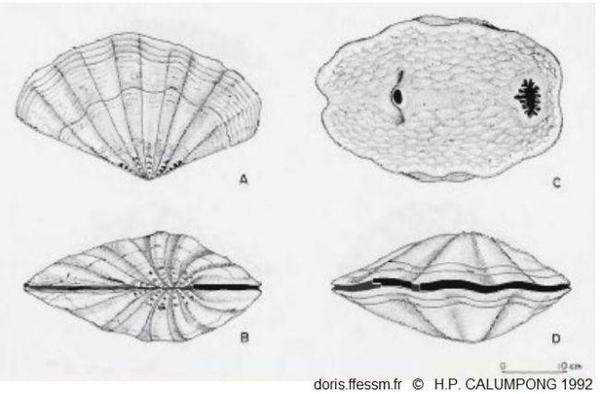
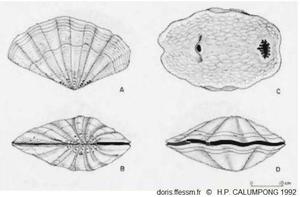
Tridacna mbalavuana dessins
A : coquille en vue latérale
B : coquille vue par la charnière
C : animal vivant vu de dessus, à droite le siphon inhalant, à gauche le siphon exhalant
D : coquille fermée
Dessins extrait de Calumpong H.P. 1992
Reproduction de documents anciens
1992


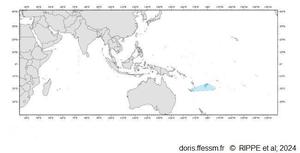
Carte de distribution
Tridacna mbalavuana a une aire de répartition réduite.
Fig 29 page 167 Rippe & al, 2024
Reproduction de documents anciens
2024
Rédacteur principal : Yves MÜLLER
Rédacteur : Cécile FAUVELOT
Vérificateur : Lucie PENIN
Responsable régional : Yves MÜLLER
Adams J., 2015, Up close with the mythical Devil clam, Tridacna mbalavuana,1p.
Calumpong H.P., 1992, The giant clam: an ocean culture manual, Australian Centre for International Agricultural Research, Monograph N°16, 68p.
Fatheree J.W., 2015, On the giant clams Tridacna mbalavuana and T. squamosina, Advanced Aquarist, 7p.
Fankboner P. V. 1981, On the siphonal eyes of giant clams (Bivalvia, Tridacnidae) and their relationship to adjacent zooxanthellae, The Veliger, 23, 245-249.
Fatherree J.W., 2002, Identifying the Tridacnid clams, Reefkeeping, 7.
Fatheree J.W., 2015, On the giant clams Tridacna mbalavuana and T. squamosina, Advanced Aquarist, 7p.
Fauvelot C., Zucco D., Borsa P., Grulois D., Magalon H., Riquet F., Andréfouët S., Berumen M.L, Sinclair-Taylor T.H., Gélin P., Behivoke F., ter Poorten J.J., Strong E.E., Bouchet P., 2020, Phylogeographical patterns and a cryptic species provide new insights into Western Indian Ocean giant clams phylogenetic relationships and colonization history, Journal of Biogeography, 47, 1086-1105.
Huelsken T., Keyse J., Liggins L., Penny S., Treml E.A., Riginos C., 2013, A novel widespread cryptic species and phylogeographic patterns within several giant clam species (Cardiidae: Tridacna) from the Indo-Pacific Ocean, PLoS ONE 8, e80858.
Ladd, H. S. (1934). Geology of Viti Levu, Fiji. Bulletin of the Bernice P. Bishop Museum 119: 1-263. page(s): 185, pl. 31 figs 2-3.
Lucas, J., Ledua, E. & Braley, R. 1991. Tridacna tevoroa Lucas, Ledua and Braley: a recently-described species of giant clam (Bivalvia, Tridacnidae) from Fiji and Tonga. Nautilus, 105(3): 92-103.
Neo M.L., Wabnitz C.C.C., Braley R.D., Heslinga G.A., Fauvelot C.,
Wynsberge S. van, Andréfouët S., Waters C., Tan A. S-H.,Gomez E.D., Costello
M.J., Todd P.A., 2017, Giant
clams (Bivalvia: Cardiidae: Tridacninae): a comprehensive update of species and
their distribution, current threats and conservation status.
Chapter 4 Oceanography and marine Biology, An Annual
Review, volume 55 in Hawkins S.J., Evans A.J., Dale A.C., Firth L.B.,
Hughes D.J., Smith J.P., 302p.
Newman W.A. & Gomez E.D. (2002). On the status of giant clams, relics of Tethys (Mollusca: Bivalvia: Tridacninae). Proceedings 9th International Coral Reef Symposium, Bali, Indonesia, 23-27(2), 1-11.
Norton J. H., Shepherd M. A.,
Long H. M., Fitt W. K., 1992, The
Zooxanthellal Tubular System in the Giant Clam, The
Biological Bulletin,183(3), 503-506.
Rippe, J.P., Young, C.N., Maison, K., Stout, C., Doss, S. 2024. Status Review Report of Seven Giant Clam Species Petitioned under the U.S. Endangered Species Act: Hippopus hippopus, H. porcellanus, Tridacna derasa, T. gigas, T. mbalavuana, T. squamosa, and T. squamosina. Draft report to the National Marine Fisheries Service, Office of Protected Resources. July 2024. 283 pp.
Tiavouane J., Fauvelot C.,2017, First record of the Devil Clam, Tridacna mbalavuana Ladd, 1934 in New Caledonia, Marine biodiversity 14, 781-782.
La page de Tridacna mbalavuana dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.





")

