

Forme ovale et allongée
Côtes rayonnantes très serrées
Stries concentriques bien marquées
Aspect quadrillé
Sinus palléal profond
Siphons séparés sur toute leur longueur
Coquille de 4 à 5 cm de longueur
Palourde commune, clovisse, palourde grise et de très nombreux noms régionaux (le nom de palourde est donné à de nombreux bivalves commercialisés).
Grooved carpet shell, cross-cut carpet shell, chequered venus, chequered carpetclam, chequered carpet shell (GB), Vongolo nera, cappa incrocicchiata (I), Teppichmuschel, Kreuzmuster Teppichmuschel, Gekreuzte Teppichmuschel, Gegitterte Venusmuschel (D), Tapijtschelp, getraliede tapijschelp, geruite tapijschelp (NL), Almeja fina (E), Amejoa (P).
Venus decussata Linnaeus, 1758
Paphia decussata (Linnaeus, 1758)
Tapes decussata (Linnaeus, 1758)
Tapes decussatus (Linnaeus, 1758)
Tapes (Ruditapes) decussatus (Linnaeus, 1758)
Venerupis decussata (Linnaeus, 1758)
Cuneus reticulatus da Costa, 1778
Venus fusca Gmelin, 1791
Venus obscura Gmelin, 1791
Venus variegata Gmelin, 1791
Venus florida Poli, 1795
Venus truncata Lamarck, 1818
Tapes decussatus var. quadrangula Jeffreys, 1864
Venus vesta Brusina, 1870
Tapes extensus Locard, 1886
Tapes decussatus var. albida Bucquoy, Dautzenberg & Dollfus, 1893
Tapes decussatus var. intermedia Bucquoy, Dautzenberg & Dollfus, 1893
Tapes decussatus var. major Bucquoy, Dautzenberg & Dollfus, 1893
Tapes decussatus var. radiata Bucquoy, Dautzenberg & Dollfus, 1893
Tapes decussatus var. texta Bucquoy, Dautzenberg & Dollfus, 1893
Tapes decussatus var. varians Bucquoy, Dautzenberg & Dollfus, 1893
Atlantique Nord-Est, Manche, Méditerranée, mer Rouge
Zones DORIS : ● Europe (côtes françaises), ○ [Méditerranée française], ○ [Atlantique Nord-Est, Manche et mer du Nord françaises], ○ [Mer Rouge]Ruditapes decussatus est présente en mer du Nord, en Manche, en Atlantique Est jusqu'au golfe de Guinée (Congo) ainsi qu'en Méditerranée.
Depuis l'ouverture du canal de Suez on la retrouve au nord de la mer Rouge.
Ruditapes decussatus vit enfouie à quelques centimètres (7 cm, maximum 15 cm) de profondeur dans le substrat* dans l'étage infralittoral*. Elle apprécie des substrats variés de sable, de petits graviers vaseux et de vase et particulièrement les zones côtières abritées comme les lagunes d'eaux saumâtres (les étangs de la côte méditerranéenne) communiquant avec la mer, les rias, les estuaires.
Cette espèce est présente depuis la mi-marée jusqu’à des profondeurs moyennes de 1 à 3 m mais rarement au-delà de 10 m. Le pied* puissant lui permet de s'enfouir rapidement et de se tenir dans le sédiment. Les limites écologiques sont comprises entre 5 °C et 30 °C pour la température, et de 15 à 40 ‰ (ou psu) pour la salinité. Ruditapes decussatus est donc une espèce eurytherme* et euryhaline*.
Comme de nombreux bivalves fouisseurs, sa distribution se fait par taches.
Les gisements peuvent présenter d’exceptionnelles densités dans les lagunes saumâtres communiquant avec la mer (bassin d’Arcachon, lagunes de Méditerranée).
Ruditapes decussatus possède deux valves* équivalves* et inéquilatérales* qui forment un ensemble ovale légèrement allongé à quadrangulaire et aplati dans la région postérieure. Sur les valves, des côtes rayonnantes très serrées ainsi que des stries concentriques assez prononcées donnent un aspect quadrillé. Cet aspect est plus marqué dans les parties postérieures et antérieures de la coquille. La marge inférieure est lisse.
La couleur externe peut varier de blanchâtre à grisâtre avec des nuances de brun, elle est rarement unie, le plus souvent avec des motifs extrêmement variables de taches, points, lignes ou chevrons bruns à noirs, souvent disposés en bandes axiales, principalement dans la zone postérieure qui est plus sombre. Les motifs des deux valves peuvent différer.
Le périostracum* est très fin et mat. Le ligament* est externe.
Chaque valve possède trois dents cardinales* : celle du centre dans la valve gauche et celles centrale et postérieure de la valve droite sont bifides. Il n’y a pas de dents latérales. La ligne palléale* et les cicatrices des muscles adducteurs* sont visibles. Le sinus* palléal est profond, en forme de U sans toutefois dépasser la ligne médiane des valves. Les siphons* sont séparés sur toute leur longueur. La partie inférieure du sinus est distincte de la ligne palléale sur toute sa longueur.
Les surfaces internes, blanches et brillantes, sont souvent teintées de jaune ou d'orange et d'une couleur bleuâtre le long du bord dorsal.
La taille moyenne est comprise entre 4 et 5 cm de longueur, cependant des individus dépassant les 8 cm ont déjà été observés.
Les différentes espèces de palourdes présentent une grande variabilité d’aspect. Il y a donc un risque de confusion avec d'autres espèces.
Ruditapes philippinarum : la palourde japonaise ou philippine est morphologiquement et écologiquement très proche de R. decussatus (cette dernière est cependant un peu plus exigeante). Les côtes rayonnantes sont plus marquées et plus écartées, les stries concentriques sont plus espacées. Le sinus* palléal* est moins profond et plus arrondi. Les siphons* sont soudés sur les ¾ de leur longueur. La couleur de la coquille est très proche. C’est la seule palourde que l’on peut trouver dans la vase molle. R. philippinarum (originaire de l'océan Pacifique) est probablement cosmopolite en mers tempérées et subtropicales. Cette espèce est présente de l’ouest de la Norvège jusqu’en Méditerranée occidentale.
Polititapes aureus (Gmelin, 1791) ou palourde jaune. Elle a une forme plus allongée dans la largeur du côté postérieur que R. decussatus. Les stries sur les valves sont marquées mais très légèrement. Sa couleur externe est nuancée de blanc crème, d'ocre et de brun avec parfois des motifs. La couleur interne tend vers un jaune or caractéristique à l'espèce. La zone de répartition de Polititapes aureus va de la Norvège au nord jusqu’en Galice (Espagne) au sud. Elle est également présente dans toute la Méditerranée, mer Noire comprise, ainsi que dans l’archipel des Açores à l’ouest de l’Afrique. Cette palourde préfère les zones calmes de l'infralittoral*. Elle vit enfouie dans des sédiments vaseux ou sablo-vaseux, voire de graviers, du médiolittoral* inférieur jusqu’à une quarantaine de mètres de profondeur.
Venerupis corrugata (Gmelin,1791) : la palourde poulette possède une coquille de forme ovale allongé, son sinus palléal arrondi atteint ou dépasse la ligne médiane des valves* et est séparé par un espace étroit de la ligne palléale. Elle mesure 4 à 5 cm de longueur. Sa couleur est plutôt jaune orangé avec parfois un peu de blanc. Elle peut s'enfoncer dans le sédiment jusqu'à 5 cm de profondeur et on la trouve jusqu'à 35 m de profondeur, sur des fonds de sable fin ou sablo-vaseux, dans les endroits très abrités comme les baies, les golfes ou les estuaires. C'est à mi-hauteur de l'estran* que ce coquillage est le plus abondant. Cette espèce est présente en Atlantique des côtes de Norvège à l'Afrique du Sud, en Manche et en Méditerranée.
Polititapes rhomboides (Pennant, 1777) aussi appelée palourde rose, elle a une forme proche de R. decussatus. Cependant la palourde rose est plus allongée et plus ovale, d'où le nom latin [rhomboïde]. Autre caractéristique, ses stries concentriques sont beaucoup plus marquées. Sa coloration est nuancée de blanc crème, de brun, de rouge orangé et de rose avec parfois des motifs.
Cette palourde est très commune en mer du Nord, en Manche et en Atlantique Nord-Est où elle est recensée de la Norvège au nord jusqu’aux côtes marocaines au sud. Elle est plus rare en Méditerranée.
La palourde rose vit enfouie dans le sédiment. Ubiquiste*, on peut la trouver aussi bien dans les sables grossiers plus ou moins vaseux, le sable coquillier, les fonds de graviers ou le faciès* du maërl*. Elle est présente de l’étage infralittoral* inférieur jusqu’à 180 m de profondeur environ, c’est cependant dans les 50 premiers mètres que sa densité est la plus importante.
Ruditapes decussatus est un suspensivore* filtreur* opportuniste. Cette espèce se nourrit et respire grâce aux siphons* qui affleurent à la surface du sédiment. L'eau entre par le siphon* inhalant*, elle est filtrée par les branchies* qui retiennent les particules alimentaires (et l’oxygène). L'eau est alors rejetée par le siphon exhalant*. Le régime alimentaire est composé de phytoplancton* et de matières organiques en suspension.
Chez Ruditapes decussatus comme chez les autres palourdes, les sexes sont séparés (espèces gonochoriques*), mais il n’y a pas de dimorphisme sexuel*.
La première maturation sexuelle est obtenue dès la première année (lorsqu'elle mesure environ 20 mm de longueur). La période de reproduction s'étend de juin à septembre.
Le cycle de reproduction débute lorsque la température de l’eau avoisine les 12 à 20 °C. Après un mois de maturation, la ponte est déclenchée. Les géniteurs expulsent leurs gamètes* (ovules* ou spermatozoïdes*) par le siphon* exhalant*. Une femelle peut émettre jusqu'à 3 millions d'ovules. La fécondation* a lieu en pleine eau.
Lorsque les conditions sont réunies, il peut y avoir deux pontes au cours de la même saison, une à la fin du printemps et l'autre à la fin de l'été.
Après une courte vie planctonique* (larves* trochophores* puis véligères* planctotrophiques* pendant 2 à 4 semaines), les larves (longueur de coquille : 190 à 235 µm) se posent sur le fond et adoptent une vie benthique*. A ce stade, la jeune palourde mesure 0,5 mm.
La croissance dépendrait de la température moyenne et de la disponibilité de la nourriture.
De nombreux endoparasites* sont présents au stade larvaire : Bacciger bacciger (Rudolphi, 1819) Nicoll, 1914, Prosorhynchoides haimeana (Lacaze-Duthiers, 1854), Bucephalus labracis Paggi & Orecchia, 1965, Gymnophallus minutus (Cobbold, 1859), Cercaria ophicerca Palombi, 1934, Gymnophallus fossarum Bartoli, 1965, Gymnophallus megacoela Palombi, 1934, Gymnophallus rostratus Bartoli, 1982, Gymnophallus strigatus (Lebour, 1908) Palombi, 1940, Himasthla ambigua Palombi, 1934, Himasthla quissetensis (Miller & Northup, 1926) Stunkard, 1934, Lasiotocus longicystis Bartoli, 1965, Lepocreadium album (Stossich, 1890) Stossich, 1904, Metacercaria acherusiae Palombi, 1934,
Et d’autres sont présents au stade adulte : Conchyliurus cardii Gooding, 1957, Herrmannella rostrata Canu, 1891, Mytilicola intestinalis Steuer, 1902
Les palourdes ont la capacité de se déplacer dans le substrat.
La croissance est influencée par deux facteurs principaux, la température et la richesse alimentaire. Plus la température est élevée, plus la filtration est importante. L'optimal de croissance a été observé dans une eau à 20 °C pour une salinité de 30 ‰ (ou psu). En période hivernale, la croissance est bien souvent arrêtée.
Lorsque le milieu offre de bonnes conditions, on observe d'importantes colonies de palourdes.
Le régime alimentaire de beaucoup de poissons est à base de mollusques. Ainsi la daurade royale (Sparus aurata) et le baliste (Balistes capriscus) exercent une forte prédation sur la palourde.
La palourde croisée européenne fait l'objet d'une exploitation commerciale depuis des décennies. Elle est particulièrement prisée en Espagne et en Italie.
Elle est pêchée en apnée ou à l'aide d'une drague tirée à la main ou avec un bateau, d'un râteau, ou encore à la main.
Etant donnée la bonne valeur commerciale du coquillage, beaucoup d'efforts ont été portés sur la vénériculture (culture de la palourde). Il faut entre 24 et 36 mois pour produire une palourde commercialisable. Cette espèce est consommée fraîche ou en conserve (sa chair crue, très fine, est plus tendre que celle de la praire commune Venus verrucosa).
Toutefois, elle a été remplacée, dans les élevages, par une espèce apparentée R. philippinarum moins exigeante.
Comme de nombreux bivalves filtreurs, cette espèce peut jouer le rôle de bioindicateur de la qualité du milieu. Elle est souvent contaminée par divers éléments traces métalliques (les métaux lourds) et les microplastiques. Du fait de sa valeur commerciale, elle fait l’objet de nombreuses recherches.
Ces espèces font l'objet d'une réglementation sur la taille minimale de capture.
En Manche, la taille minimale doit être supérieure à 4 cm (dans la limite de cent individus par jour maximum) pour la palourde croisée européenne selon l'arrêté 55/2007 réglementant l'exercice de la pêche maritime à pied, à la nage ou sous-marine en Manche.
En Méditerranée, la taille minimale doit être supérieure à 3,5 cm pour la palourde croisée européenne selon l'arrêté du 19 mars 2007 déterminant la taille minimale et le poids minimal de capture et de débarquement des poissons et autres organismes marins.
La réglementation européenne est moins restrictive : la taille minimale de pêche est de 25 mm pour Ruditapes spp. dans le Règlement (CE) n° 1967/2006 du Conseil du 21 décembre 2006 concernant des mesures de gestion pour l'exploitation durable des ressources halieutiques* en Méditerranée.
Ne pas hésiter à se renseigner auprès des Affaires Maritimes du département pour des informations complémentaires à la réglementation.
Le mot palourde est apparu dans la langue française au 16e siècle. Il vient du latin [peloris] qui désigne un gros coquillage.
Ruditapes vient du latin [rudis] = brut, qui est à l'état naturel, non façonné, rude, âpre, dur ; et du latin [tapes] = tapis, tapisserie. Ce nom de genre a été créé par le naturaliste italien Alessandro Chiamenti (1848-19 ??) en 1900 ; à partir du nom de genre Tapes créé, en 1811, par le malacologiste autrichien Johann Karl Megerle von Mülhfeld (1765-1840) pour des coquilles dont l'aspect rappelle les motifs géométriques de certains tapis ou paillassons.
decussatus est le participe passé du mot latin [decusso] = croisé, en forme de croix du fait des stries concentriques et rayonnantes. Ce nom d’espèce a été donné, en 1758, par le naturaliste suédois Carl von Linné (1707-1778).
Numéro d'entrée WoRMS : 231749
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Bivalvia / Lamellibranchia / Pelecypoda | Bivalves / Lamellibranches / Pélécypodes | Mollusques aquatiques, filtreurs, au corps comprimé latéralement. Coquille composée de 2 valves articulées disposées de part et d’autre du plan de symétrie. Absence de tête, de pharynx, de radula et de glande salivaire. |
| Sous-classe | Autobranchia | Autobranches | |
| Infra-classe | Heteroconchia | Hétéroconchie | |
| Subter-classe | Euheterodonta | Euhétérodonte | |
| Super ordre | Imparidentia | Imparidenties | |
| Ordre | Venerida | Vénérides | Coquille mince, allongée, dure et costulée, bâillante à une ou aux deux extrémités. Ligament à la fois interne et externe. |
| Super-famille | Veneroidea | ||
| Famille | Veneridae | Vénéridés | Coquille équivalve pour la plupart des espèces. De forme ronde, ovale ou encore oblongue. Ecusson distinct, présence de stries concentriques et parfois d’éléments rayonnants |
| Sous-famille | Tapetinae | ||
| Genre | Ruditapes | ||
| Espèce | decussatus |
") Bivalves (ou Lamellibranches)
Bivalves (ou Lamellibranches)

Palourde croisée d'Europe
On remarque les stries rayonnantes très rapprochées, ainsi que quelques stries concentriques.
Bassin d’Arcachon (33)
10/08/2008
Bivalves (ou Lamellibranches)

Détail des siphons
La palourde croisée d'Europe à des siphons séparés sur toute la longueur, contrairement à la palourde japonaise qui a ses siphons soudés jusqu’aux ¾.
Petit jeu : identifier les coques (coquille striée et siphons jaunes et courts) mélangées aux palourdes dans le panier ...
Trébeurden (22)
06/08/2008

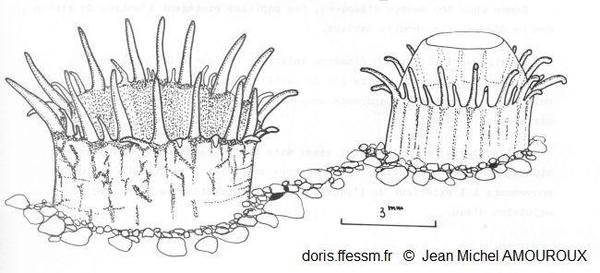
Détail des siphons
Les siphons* sont séparés sur toute leur longueur d’une longueur égale à celle du corps et affleurent la surface du sédiment. Le siphon inhalant*, de section légèrement ovale, possède de grandes papilles* inégales orientées vers l’intérieur ou l’extérieur et de plus petites papilles orientées vers l’intérieur du siphon. Le siphon exhalant* a un diamètre de moitié inférieur à celui de l’inhalant et possède une rangée de papilles égales et une valvule très fine.
fig 18 page 57 et 58, J.M. Amouroux 1980.
Reproduction de documents anciens
1980


Palourde croisée d'Europe enfouie dans le sédiment
Les trous sont espacés d’environ la longueur de la coquille caractéristique de la palourde européenne.
Bassin d’Arcachon (33)
10/08/2008

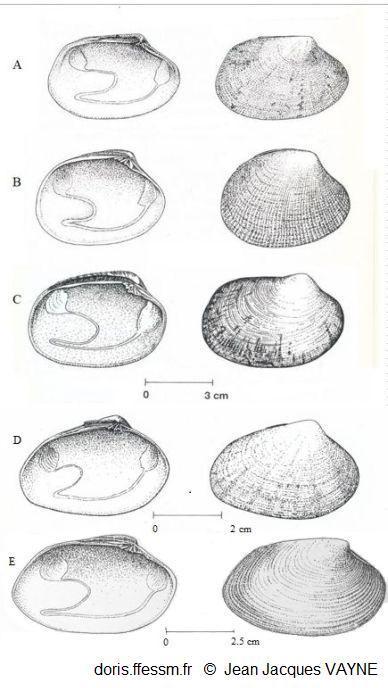
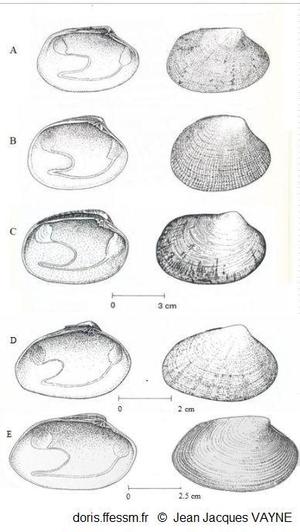
Comparaison de cinq palourdes
à gauche vue intérieure de la valve gauche, à droite vue extérieure de la valve droite
A: Ruditapes decussatus
B: Ruditapes philippinarum
c: Venerupis corrugata
D :Polititapes aureus
E: Polititapes rhomboides
D’après les dessins de J.J. Vayne in Quéro et al. 1998 (pp : 121, 123, 145, 146, 126)
Reproduction de documents anciens
1998


Palourde japonaise et palourde croisée d'Europe
Comparaison entre la palourde japonaise Ruditapes philippinarum (à gauche) et la palourde croisée d'Europe (à droite).
Bassin d’Arcachon (33)
10/08/2008


Une inconsciente !
Cette palourde restait visible sur l'estran* : un promeneur en a fait son apéritif impromptu !
Trégastel (22), estran
09/05/2009


Gravure ancienne
Sur cette gravure ancienne, on distingue le pied et les deux siphons* de notre palourde
Fig. 2, planche 83 dans "Histoire naturelle des mollusques" par G.P. Deshayes
Reproduction de documents anciens
1848


Dessin ancien par A. D’Apreval
L’auteur nous montre en haut la forme typique (166) et en bas la coloration plus foncée de la var. fusca (165).
Fig. 165 & 166, planche 50 dans "Atlas de poche des coquilles des côtes de France" par Ph. Dautzenberg
Reproduction de documents anciens
1913
Rédacteur principal : Cédric CONTI
Vérificateur : Philippe LE GRANCHÉ
Responsable historique : Véronique LAMARE
Responsable régional : Yves MÜLLER
Amouroux J. M., 1980, Etude monographique des siphons de quelques mollusques bivalves : adaptation et morphologie, Océanis, 5(1), 33-89, Institut Océanographique, Paris.
Ben Ouada H., Mehioub M.N., Medhioub A., Jammoussi H., Beji M.,1997, Variabilité morphométrique de la palourde Ruditapes decussatus (Linné, 1758) le long des côtes tunisiennes, Haliotis, 27, 43-55.
Bodoy A., Plante-Cuny M.R.,1984, Relations entre l'évolution saisonnière des populations de palourdes (Ruditapes decussatus) et celle des microphytes benthiques et planctoniques (Golfe de Fos- France), Haliotis,14, 71.
Delgado M., Camacho A.P., 2005, Histological study of the gonadal development of Ruditapes decussatus (L.) (Mollusca: Bivalvia) and its relationship with available food*, Scientia marina, 69(1), 87-97.
Garcia F., 1993, Interprétation des stries valvaires pour l'évaluation de la croissance de Ruditapes decussatus L., Oceanologica Acta, 1, 199-203.
Gérard A., 1978, Recherche sur la variabilité de diverses populations de Ruditapes decussatus et Ruditapes philippinarum (Veneridae, Bivalvia). Thèse 3ème cycle. Univ. Bretagne Occidentale, Brest, France, 149p.
His E., Cantin C., 1995, Biologie, physiologie des coquillages, IFREMER, 118p.
Kamara A., Rharbi N., Ramdani M., Berraho A., 2008, Recherches préliminaires au développement de l'élevage de la palourde européenne (Ruditapes decussatus L.) sur les côtes marocaines et au repeuplement des sites surexploités, Bulletin de la Société Zoologique de France,133(1-3), 193-202.
Locard A., 1886, PRODROME DE MALACOLOGIE FRANÇAISE, CATALOGUE GENERAL DES MOLLUSQUES VIVANTS DE FRANCE. MOLLUSQUES MARINS. Lyon: H. Georg & Paris, Baillière, + 778 p.
Nalbone L., Giarratana F., Genovese M., Panebianco A.,2024, Occurrence of microplastics in store-bought fresh and processed clams in Italy, Marine Pollution Bulletin, 206, 24p.
Rodriguez-Moscoso E, Arnaiz R., 1998, Gametogenesis and energy storage in a population of the grooved carpet-shell clam, Tapes decussatus (Linne, 1787), in northwest Spain, Aquaculture,162(1-2), 125-139.
Smaoui-Damak W., Berthet B., Rebai T., Hamza-Chaffai A., 2008, Effets de la pollution par les métaux sur le potentiel reproducteur de la palourde Ruditapes decussatus (L.) issue du Golfe de Gabès (Tunisie), Bulletin de la Société Zoologique de France, 133(1-3), 131-140.
Trigui El Menif N., Le Pennec M., El Abed A., 2000, Variabilité génétique de la palourde Ruditapes decussatus de la côte nord-est tunisienne, Haliotis, 30, 13-22.
Trigui El Menif N., Le Pennec M., Maamouri F., Medhioub K., 1998, La bioaccumulation de métaux chez le vénéridé Ruditapes decussatus (L.) récolté sur le littoral tunisien, Haliotis, 28, 21-30.
Xie Q., Burnell GM.,1994, A comparative study of the gametogenic cycles of the clams Tapes philippinarum (A. Adams & Reeve 1850) and Tapes decussatus (Linnaeus) on the south coast of Ireland. Journal of Shellfish Research, 13(2), 467-472.
La page de Ruditapes decussatus dans l'Inventaire National du Patrimoine Naturel : INPN
La page de Ruditapes decussatus sur le site de référence de DORIS pour les mollusques :MolluscaBase
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.










")

