

Corps tubulaire relativement fin
Couleur uniformément noire à reflets brun-rouge possibles
Tégument lisse, papilles courtes et peu nombreuses
Bouche ventrale, anus terminal
Canaux de Cuvier longs et fins
Holothurie noire, trépang à canaux blancs, holothurie à filaments blancs
Shy black sea cucumber, white threads fish, black trepang, black dog (GB), Weißgefleckte Seegurke (D), Sisigo (Samoa), Ka la lee (Birmanie), Patola (Malaisie), Lolosong (langue Bajo, Indonésie)
Stichopus leucospilota Brandt, 1835
Holothuria vagabunda Selenka, 1867
Holothuria lagoena Haacke, 1880
Indo-Pacifique tropical, mer Rouge
Zones DORIS : ● Indo-Pacifique, ○ [Mer Rouge]On rencontre cette espèce dans l'océan Indien (très présente à la Réunion, elle semble peu représentée sur Mayotte) ainsi que dans l'océan Pacifique tropical Est et centre jusqu'à Hawaï.
L'espèce est présente également en mer Rouge et dans le golfe Persique.
On trouve cette holothurie par petits fonds (de 0 à 10 mètres) dans les zones sableuses des récifs, notamment là où se trouvent de nombreux blocs détritiques. Elle apprécie les zones calmes.
Il s'agit d'une holothurie de taille moyenne à grande (maxima de 60 cm de longueur et 6 cm de large) au corps tubulaire relativement fin s'amincissant en partie antérieure. La section est approximativement circulaire.
La couleur est uniformément noire, souvent avec des reflets brun-rouge plus ou moins prononcés. Le tégument, lisse et peu épais, est fréquemment couvert d'une mince couche de sable fixée par du mucus. On peut parfois observer également une desquamation de ce tégument.
Le bivium* (face dorsale) porte des papilles coniques courtes et de fins podia* (pieds ambulacraires*) à distribution irrégulière hors aires ambulacraires.
Le trivium* (face ventrale) est légèrement aplati et présente de nombreux podia courts et larges, s'achevant par une ventouse blanche et répartis sur 4 ou 5 rangs sur les radius*. Les podia sont moins nombreux et sont dispersés dans les interradius*.
La bouche est ventrale (positionnée au plus près du substrat) et entourée par 20 tentacules assez longs.
L'anus est terminal.
Les tubes de Cuvier, facilement expulsés, sont nombreux, longs et fins, de couleur blanc bleuté.
Holothuria atra peut être confondue dans son biotope avec Holothuria leucospilota, d'autant qu'elles se trouvent souvent sur les mêmes sites. Holothuria atra est cependant plus petite et le plus souvent couverte d'une couche de sable sauf en deux rangées de zones circulaires plus ou moins parallèles, laissant apparaître un tégument noir. D'autre part, ses tentacules sont courts et elle ne possède pas d'organe de Cuvier.
Actinopyga miliaris (qui peut aussi porter le nom vernaculaire d'holothurie noire) est plus petite, de forme ovoïde et possède des dents anales.
Holothuria coluber : le tégument est gris-bleu foncé couvert de petits points clairs, le corps est plus large en partie antérieure, les papilles sont jaunes et placées sur des verrucosités. Les tentacules sont de couleur crème.
C'est une espèce détritivore qui ingère le sable, n'en retenant que les particules organiques végétales et animales qui y sont contenues. Elle rejette ensuite ses fèces* en une série de petits tronçons cylindriques. L'holothurie participe ainsi de façon notable, tant par élimination de débris organiques que par remaniement continuel du substrat, à l'équilibre des écosystèmes côtiers.
La reproduction peut être sexuée : mâles et femelles se dressent le plus haut possible pour diffuser largement des gamètes qu'ils émettent alors. On les trouve ainsi parfois au sommet des coraux. L'observation peut en être faite le soir et le plus souvent de nuit.
L'émission des mâles est beige, celle des femelles est rose. La fécondation se fait dans la colonne d'eau.
Les quatre stades larvaires (gastrula, auricularia, doliolaria, pentactula) se déroulent en pleine eau ; 33 jours après la fécondation et définitivement posé sur le substrat, le juvénile d'un millimètre va évoluer vers le stade adulte.
La reproduction peut également être asexuée et se produire par scissiparité*. Les individus en cours de scission cessent de se nourrir et se cachent pour se protéger.
L'endroit du corps où a lieu la scission est propre à chaque espèce ! Ainsi, pour H. leucospilota, la scission se fait au cinquième de la longueur du corps, une partie du système respiratoire restant dans chacun des éléments.
Les parties postérieures semblent avoir de meilleures chances de survie que les antérieures pendant le processus de régénération. La régénération peut commencer dès le dernier stade de la scission et dure plusieurs semaines.
Ce mode de reproduction peut coexister avec le premier, ou ne pas se produire dans certaines populations, probablement en fonction des conditions environnementales.
Holothuria leucospilota est parfois parasitée par des Eulimidés (Gastéropodes), ainsi que par un poisson Carapidé, Encheliophis vermicularis.
Espèce très commune qu'on trouve parfois en agrégations importantes.
Les spicules* présents dans le tégument sont en tourelles et boutons sur l'ensemble du corps, s'y ajoutent des plaques sur les podia ventraux et des bâtonnets sur les podia dorsaux. Les tentacules ne sont pas équipés de spicules.
La défense de l'holothurie noire de l'Indo-Pacifique est assurée par l'organe de Cuvier, qui se trouve près du cloaque, les tubes collants étant projetés par l'anus. La pression d'eau permettant leur expulsion les fait alors s'allonger et se disperser autour de l'agresseur qui s'y empêtre. Ces tubes contiennent une substance toxique, l'holothurine, susceptible de provoquer chez l'homme des irritations. La médecine rapporte également des complications plus graves.
L'éviscération est enfin le dernier recours utilisé par l'holothurie, donnant ainsi satisfaction au prédateur, qui laissera s'éloigner l'animal, au prix du temps de vulnérabilité requis par la régénération de ses organes.
On assiste aussi à des moments de desquamation générale, comme si l'holothurie muait : J. P. McMurrich a suggéré en 1894 un lien avec l'excrétion de résidus de l'activité métabolique par les arbres respiratoires, résidus qui seraient portés vers le tégument et ainsi périodiquement éliminés. Il peut également s'agir de déparasitage externe. On peut voir parfois des poissons picorer cette paroi dont se débarrasse l'holothurie.
Une étude menée sur Holothuria forskali a montré que cette espèce, comme ses cousins Echinodermes les oursins et les étoiles de mer, possède sur chaque disque podial un système cellulaire complexe permettant l'adhérence au substrat et donc la locomotion. L'épiderme du disque podial est composé de 5 types de cellules. Deux d'entre eux produisent un mucus différent qui, par mélange, devient collant et permet l'adhésion. Un troisième type cellulaire, composé de cellules ciliées dites sécrétrices, permet selon toute vraisemblance de dissoudre la couche superficielle de « colle » fabriquée à la surface du disque et de provoquer le détachement. Les deux derniers types de cellules sont composés de cellules ciliées non-sécrétrices et de cellules de soutien. La stimulation des deux types de cellules ciliées coordonnerait les sécrétions d'adhésion et de détachement, et permettrait ainsi le déplacement. Il est probable que cette organisation du disque podial se retrouve de manière plus ou moins bien conservée chez toutes les espèces d'holothuries, fonction de la modification des podias selon les espèces. La détermination de la nature des sécrétions produites par ces classes d'Echinodermes pourrait à terme déboucher sur la fabrication de colles biomimétiques efficaces en milieu humide ainsi que d'antifouling pour l'entretien des bateaux.
Holothuria leucospilota est parfois consommée à une échelle locale, mais moins que les Holothuria nobilis, scabra et fuscogilva, par exemple. Elle fait d'ailleurs partie des espèces les moins chères, parce que les moins recherchées.
Ses propriétés médicinales sont étudiées par les laboratoires.
Dans [Chen 2003], on peut lire que les holothuries en général sont connues depuis très longtemps en Asie pour leurs propriétés médicinales. Du point de vue médical moderne, les comprimés ou gélules produits à partir de certaines holothuries sont considérés comme un supplément alimentaire particulièrement riche en protéines, qui sont aussi capables de diminuer les douleurs articulaires. Les composants de la bêche de mer sont encore connus pour ralentir la progression des infections virales (le sulfate de chondroitine notamment est utilisé par les Japonais dans le cadre d'une thérapie anti-VIH) et pour des propriétés anti-cancérigènes (selon des études chinoises).
Holothuria leucospilota est une espèce de faible valeur commerciale ; elle n'est pas mentionnée en annexe de la CITES (La Convention sur le commerce international des espèces de faune et de flore sauvages menacées d'extinction).
Néanmoins, de nombreux pays réglementent la pêche des holothuries en général par fixation de quotas, de saisons de pêche, de limitations des aires, de permis divers ou de tailles minimales.
Holothurie noire de l'Indo-Pacifique est une proposition du site DORIS, apportant la précision géographique au nom vernaculaire francophone localement rencontré d'holothurie noire. Cela permet de distinguer cette espèce d'Holothuria forskali et d'Actynopyga miliaris (toutes deux appelées également holothuries noires !).
L'un des autres noms vernaculaires (holothurie à filaments blancs) évoque les canaux de Cuvier.
Holothuria : Le terme grec [holothourion] est le nom donné par Aristote à un animal que les ambiguïtés de sa description rendent impossible à déterminer. Le mot latin [holothuria] a désigné d'abord des cnidaires, puis a été appliqué aux échinodermes par Linné.
Une tentative de décomposition du mot grec holothourion en [holo-] = entier ; et [thouro-] = impétueux, produit l'interprétation fréquente : « tout à fait impudique ». La forme phallique de ces animaux motive cette interprétation.
leucospilota : du grec [leukos] = de couleur blanche ; et [spilos] = tache sur la peau, bouton.
NB : le nom scientifique complet de cette espèce est Holothuria (Mertensothuria) leucospilota. Le sous-genre Mertensiothuria a été créé par Deichmann en 1958. La descriptrice ne donne pas les raisons de son choix pour le nom du sous-genre, mais il semble évident qu'il est donné en l'honneur du naturaliste et botaniste allemand C. H. Mertens. Pour ce qui concerne la morphologie externe de ce sous-genre, il est caractérisé par un corps cylindrique ou à faible aplatissement du trivium, des podia et papilles dispersés sur tout le corps, l'absence de papilles anales et de collier de papilles autour des tentacules, qui sont au nombre de 18 à 20.
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Echinodermata | Echinodermes | Symétrie radiale d'ordre cinq (chez les adultes). Squelette de plaques calcaires bien développé sous le derme. Présence d'un système aquifère auquel appartiennent les podia souvent visibles extérieurement. |
| Sous-embranchement | Echinozoa | Echinozoaires | Echinodermes non étoilés de forme globuleuse ou allongée. Ce groupe renferme les oursins et les concombres de mer. |
| Classe | Holothuroidea | Holothuroïdes | Echinodermes vermiformes, ouverture buccale à l’extrémité antérieure du corps et entourée d’une couronne de tentacules rétractiles, anus postérieur, une seule gonade : holothuries, concombres de mer. Endosquelette réduit à de microscopiques ossicules ou plaques, inclus dans la paroi du corps. |
| Super ordre | Aspidochirotacea | Aspidochirotacés | |
| Ordre | Holothuriida | Holothurides | (Anciennement: Aspidochirotida / Aspidochirotes) Symétrie bilatérale, avec une sole de reptation et des podia buccaux en forme d’écusson. Présence de poumons, pas de muscle rétracteur de la bouche. |
| Famille | Holothuriidae | Holothuriidés | Podia munis d’ampoules. La gonade est placée à gauche du mésentère dorsal. |
| Genre | Holothuria (Mertensiothuria) | ||
| Espèce | leucospilota |
") Concombres de mer (Holothurides)
Concombres de mer (Holothurides)

Escalade
Escalade inhabituelle de la part d’Holothuria leucospilota : le bloc, probablement riche, a été d’abord exploité par l'échinoderme avant le retour au substrat.
Lagon de L’Ermitage, La Réunion, 1,5 m
07/08/2010
Concombres de mer (Holothurides)

Tentacules buccaux
Les tentacules buccaux palpent une pierre pour y trouver les débris organiques dont l’animal se nourrit.
Lagon de L’Ermitage, La Réunion, 1,5 m
07/08/2010


Tubes de Cuvier
Début d'une émission des tubes de Cuvier.
Lagon de L’Ermitage, La Réunion, 1,5 m
07/08/2010


Excrétion
Suite à l'ingestion, le sable se voit débarrassé des particules organiques qu’il contenait et est restitué, à l'arrière, sous la forme de petits sacs allongés.
Lagon de L’Ermitage, La Réunion, 1,5 m
04/08/2010


Emission de gamètes
Cet individu prend l’attitude nécessaire pour l’émission de gamètes.
Si la situation est plus courante en soirée et même le plus souvent de nuit, cette photo a été prise un matin. L'émission peut donc être différée.
Lagon de L’Ermitage, La Réunion, 1,5 m
18/05/2010


Petit juvénile
Ce très jeune individu mesure 2 cm. La couleur est encore claire et les podia comme les papilles sont très peu nombreux. L'individu est légèrement penché sur le côté, ce qui permet d'observer la différence entre les gros podia ventraux et les papilles dorsales, qui sont des podia transformés.
Lagon de l'Ermitage, La Réunion, 1 m, en PMT
15/02/2017


Juvénile
Cet individu de 9 cm est un juvénile. Son aspect est identique à celui des adultes.
La Réunion, 1 m
23/05/2015


Scissiparité
Un des modes de reproduction, asexué, de beaucoup d'échinodermes est la scissiparité. L'animal se divise en deux parties qui régénèreront chacune un individu complet.
Ce spécimen de 3 cm est issu d’une scission transversale dont il est la partie antérieure.
Lagon de L’Ermitage, La Réunion, 1,5 m
21/04/2010

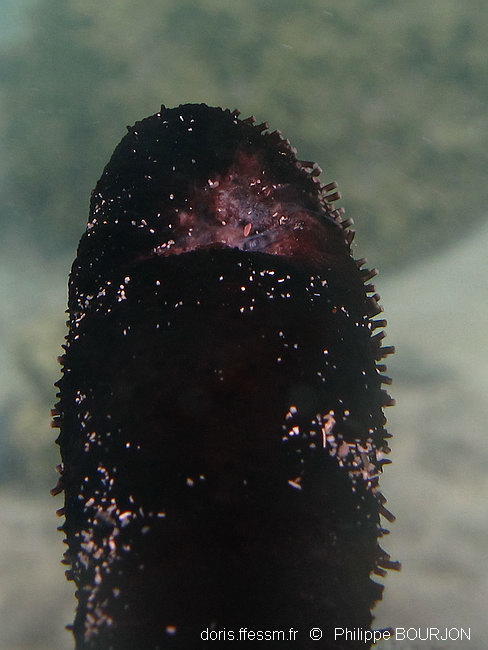
Scissiparité 2
Holothuria leucospilota peut, à des fins de reproduction, se scinder en deux parties. Voilà une des extrémités résultantes. Le point de scission n'est pas encore cicatrisé !
Ce spécimen faisait envion 6 cm et la scission devait donc avoir 1 ou 2 jours.
Lagon de l'Ermitage, La Réunion, 1,5 m
17/08/2010


Scission
Cet individu est en cours de scission comme le montre l'étranglement de son corps en partie antérieure.
La présence de gonades à côté de lui est surprenante dans la mesure où il est peu probable qu'elles lui appartiennent. Il se pourrait qu'un autre individu, agressé, se soit éviscéré non loin et que seules les gonades restent après le passage de divers prédateurs.
Lagon de l'Ermitage, La Réunion, 1,5 m
15/02/2012


Podia ventraux
Détail du trivium (partie ventrale) : les podia, courts et larges, sont pourvus d’un disque podial blanc.
Lagon de L’Ermitage, La Réunion, 1,5 m
21/05/2010


Desquamation
La desquamation de la surface du tégument est un phénomène fréquent, ayant probablement une fonction de déparasitage.
Lagon de L’Ermitage, La Réunion, 1,5 m
07/08/2010


Desquamation 2
On retrouve ce principe de desquamation chez plusieurs espèces, tropicales ou méditerranéennes. La raison exacte n'est pas encore connue avec toute la précision voulue.
Archipel des Siliman, Thaïlande
2007


Agrégation
On peut trouver sur certains sites de très nombreux spécimens d’Holothuria leucospilota sur une surface peu étendue, bien qu’elle puisse se rencontrer isolée ailleurs.
Ce comportement de rassemblement ne semble pas lié à des impératifs de reproduction.
Lagon de L’Ermitage, La Réunion, 1,5 m
07/08/2010


Gastéropodes parasites
Ces gastéropodes parasites sont des Eulimidés, qui se fixent par leur trompe dans le tégument de l'holothurie.
Ils ne présentent pas de danger réel pour leur hôte involontaire.
Lagon de L’Ermitage, La Réunion, 1,5 m
27/06/2010


En Nouvelle-Calédonie
Un individu néo-calédonien. Il est difficile de savoir ce que sont les filaments blancs visibles sur ce cliché... Restes de tubes de Cuvier ou émission de gamètes ? Le photographe a indiqué que ça semblait sortir de plusieurs endroits, au moins deux, voire trois. Ceci rend la réponse plus difficile encore.
Nouvelle-Calédonie (océan Pacifique)
11/11/2005
Rédacteur principal : Philippe BOURJON
Vérificateur : Alain-Pierre SITTLER
Correcteur : Chantal CONAND
Responsable régional : Alain-Pierre SITTLER
Chen J., 2003, Aperçu des méthodes d'aquaculture et de mariculture d'holothuries en Chine, La Bêche-de-mer, bulletin de la CPS n° 18, 18-23.
Conand C. & Mangion P., 2003, Les holothuries des récifs frangeants de La Réunion : diversité, distribution, abondance et structure des populations, La Bêche-de-mer, Bulletin de la CPS n° 17, 1-6.
Conand C., Morel C., Mussard R., 1997, Une nouvelle observation de reproduction asexuée chez les holothurie; scission dans des populations de Holothuria leucospilota à La Réunion, océan Indien, La Bêche-de-mer, Bulletin de la CPS n° 9, 5-11.
Flammang P., Jangoux M., 1992, Functionnal morphology of the locomotory podia of Holothuria forskali (Echinodermata, Holothuroida), Zoomorphology, 111(3), 167-178.
Pouget A., 2005, Abondance et distribution des holothuries présentes sur les platiers des récifs frangeants de Grande Terre à Mayotte (océan Indien), La bêche-de-mer, Bulletin de la CPS n° 21, 1-5.
Purwati P., 2005, La scissiparité chez Holoturia leucospilota dans les eaux tropicales de Darwin (Territoire du Nord de l'Australie), La Bêche-de-mer, bulletin de la CPS n° 20, 26-33.
Santos R., 2012, A multidisciplinary analysis of sea-urchin temporary adhesion: morphology, biomechanics and proteomics, 14th International Echinoderm Conference, Plenary abstract, 24p.
La page d'Holothuria (Mertensiothuria) leucospilota dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.
















")

