

Coquille triangulaire de 12 à 16 mm de longueur
Coquille presque équilatérale, nettement convexe, voire globuleuse
Extrémité postérieure avec une crête radiale caractéristique
Pas de ligament externe
Sommets, presque centraux, proéminents et nettement séparés
Dwarf surfclam, coot clam,little surf clam, duck clam (GB), Amerikaanse stranschelp (NL)
Mactra lateralis Say, 1822
Mactra rostrata R. A. Philippi, 1849
Atlantique Nord, ouest et est
Zones DORIS : ● Atlantique Nord-Ouest, ○ [Atlantique Nord-Est, Manche et mer du Nord françaises]Mulinia lateralis est présente sur les côtes orientales de l'Amérique du Nord, du golfe du Saint Laurent jusqu'au Mexique, en Colombie et aux Caraïbes.
Depuis quelques années, cette espèce est introduite en Europe,. Elle est observée en mer du Nord, de la mer des Wadden (Allemagne), dans l’estuaire de l’Escaut occidental (Pays-Bas) jusqu'au Golfe de Gascogne.
Mulinia lateralis vit peu profondément enfouie dans des sédiments mous (sable ou vase) en bas de la zone de balancement des marées, dans des estuaires, des lagunes et des eaux côtières abritées. Elle est adaptée aux variations de salinité (de 5 à 80 ‰ voire plus, elle est donc euryhaline*), de température (- 2 °C à 35 °C, elle est donc eurytherme*), à une mauvaise qualité de l’eau, à l’eutrophisation*, des conditions anoxiques* et de la pollution.
Mulinia lateralis a une coquille triangulaire de 12 à 16 mm de longueur (voire 21 mm), presque équilatérale*, nettement convexe, voire globuleuse, lisse avec de fines stries de croissance concentriques. L’extrémité antérieure de la coquille est arrondie, l’extrémité postérieure présente une crête radiale caractéristique qui donne un aspect anguleux. La surface de la coquille est blanche ou crème et un peu brillante. Le périostracum* est mince, jaunâtre ou crème.
Il n’y a pas de ligament* externe. Le ligament est interne (car logé dans le chondrophore*). Les sommets, presque centraux, sont proéminents et nettement séparés.
La surface interne est d’un blanc brillant. Le sinus* palléal* est peu profond et n’atteint pas la ligne médiane.
Le pied* est large et pointu, les siphons*, brun-clair, minces et légèrement incurvés vers le haut, sont étroitement unis et lorsqu’ils sont étendus, ils dépassent la longueur de la coquille. L’extrémité du siphon inhalant* porte 24 papilles* (alternativement longues et courtes) transparentes aux extrémités blanches, avec des points bruns entre elles. La valvule du siphon exhalant* dépasse l’extrémité des papilles.
Les spécimens juvéniles de Spisula subtruncata (da Costa, 1778) sans périostracum* peuvent être confondus avec de petits spécimens de M. lateralis, mais la présence de la crête radiale le long de l'extrémité postérieure des valves, l'aspect bulbeux et les deux sommets bien séparés, l’absence de ligament externe de M. lateralis sont décisifs pour l'identification de cette dernière.
M. lateralis préfère les eaux saumâtres avec des fonds boueux, pauvres en oxygène contrairement à S. subtruncata qui vit dans des zones sablonneuses, les deux espèces ne sont pas de véritables concurrents dans leurs habitats.
Toutefois les coquilles vides de ces deux espèces peuvent être présentes, ensemble, dans les mêmes laisses* de mer.
Les juvéniles de Mactra stultorum sont aussi délicats que ceux de M. lateralis, mais leurs contours diffèrent. La coquille de M. stultorum est plus arrondie, beaucoup moins globuleuse et surtout n'a pas la crête radiale postérieure.
L'espèce non indigène la rangia d’Atlantique (Rangia cuneata (G.B. Sowerby I, 1832)), présente en Belgique depuis 2005, est beaucoup plus grande (5 cm), plus lourde avec une coquille plus épaisse. Elle est asymétrique avec de grands sommets, des lignes concentriques épaisses et inégales, ainsi qu'une plaque charnière épaissie. Elle vit dans des eaux saumâtres ou dans des estuaires.
M. lateralis est un filtreur*. Son régime alimentaire se compose principalement de bactéries ou d'autres matières en suspension. Comme d'autres mollusques bivalves, l'animal étend ses siphons* jusqu’à la surface du sédiment, aspire l'eau par le siphon inhalant* et l'expulse par le siphon exhalant*, après avoir filtré les particules nutritives dans les branchies*.
M. lateralis peut exploiter rapidement les fortes concentrations de phytoplancton* et de seston* naturel (en particulier les bactéries) typiques des eaux eutrophes*.
Chez Mulinia lateralis, les sexes sont séparés. Les adultes matures (à une longueur inférieure à 1 cm pour les femelles) peuvent se reproduire plus d’une fois par an (voire toute l’année au Mexique).
La femelle produit jusqu'à 2 millions d'ovules (roses à rouges voire orange) à la fois. Le sperme* des mâles est blanc. Les gamètes* sont libérés dans l’eau de mer. La fécondation* est externe. Rapidement des larves* trochophores* planctotrophes* éclosent, puis se transforment en véligères*. Ces larves se dispersent dans le zooplancton* et mettent de 1 à 3 semaines à se développer jusqu'au stade pédivéligère* (véligère munie d’un pied) et ainsi se disperser sur d’assez grandes distances. Ensuite elles rejoignent le fond et se métamorphosent. Les juvéniles grandissent rapidement et atteignent la maturité sexuelle en l'espace de 2 mois.
Le temps de génération est très court et ainsi cette espèce peut avoir de nombreux descendants et coloniser de nouveaux espaces..
la durée de vie est d'environ deux ans.
Les populations de Mulinia lateralis peuvent être abondantes, de 6 000 à 21 000 individus par m².
Mulinia lateralis peut héberger des endoparasites* au stade larvaire* comme des Cercaria apalachiensis Holliman, 1961, Cercaria granosa Holliman, 1961 ou Cercaria imbecilla Holliman, 1961.
Dans son aire de répartition indigène, M. lateralis est un hôte du pathogène parasite Perkinsus chesapeaki (l'infection s'appelle la perkinose). Ce protozoaire, présent également chez Mya arenaria affecte probablement sa croissance, sa reproduction et peut-être sa mortalité et donc son exploitation commerciale.
P. chesapeaki a été récemment signalé sur les côtes méditerranéennes et atlantiques de France et d'Espagne, infectant des palourdes et des coques, notamment Ruditapes decussatus, R. philippinarum et Cerastoderma edule (espèces également exploitées commercialement), et affectant éventuellement la production.
L'animal est très actif dans ses mouvements, le pied se déplaçant avec une grande vigueur.
Mulinia lateralis est une espèce opportuniste qui croît et se reproduit rapidement.
Elle colonise rapidement les sites perturbés, par exemple lorsque la concurrence est réduite par un événement catastrophique tel qu'une inondation ou une sécheresse (provoquant des salinités extrêmes), des incidents de pollution ou une mauvaise qualité de l'eau et de faibles niveaux d'oxygène. Elle peut rapidement devenir dominante dans les zones où les conditions lui sont optimales.
Sur les côtes atlantiques ouest, de nombreux organismes se nourrissent de Mulinia lateralis comme l'étoile de mer grise (Luidia clathrata qui s'en nourrit de manière sélective) les gastéropodes de la famille des Naticidés ainsi que le busycon à nodules Busycon carica.
Mulinia lateralis vit très près de la surface des sédiments et possède une coquille fine, elle est donc facilement accessible aux prédateurs tels que les crabes, les poissons, les échassiers, les gastéropodes, les étoiles de mer, les canards et les oiseaux de rivage.
La première observation en 2017, dans la mer des Wadden (Allemagne) a été suivie quelques mois après dans l’estuaire de l’Escaut occidental (Pays-Bas), puis sur la côte belge en 2019, sur les côtes britanniques en 2021 et enfin dans le golfe de Gascogne en 2024.
Sa capacité à coloniser rapidement les zones perturbées, sa fécondité élevée, son temps de génération court, sa tolérance à l'anoxie*, aux variations de salinité et aux températures extrêmes, ainsi que sa capacité à former des colonies très denses peuvent l'amener à entrer en compétition avec les espèces indigènes pour la nourriture et l'espace. L'état de conservation de nombreux sites pourrait être dégradé si M. lateralis y établissait des populations denses. Elle peut avoir un impact sur la production commerciale de coquillages, soit par la concurrence, soit par l'introduction du pathogène parasite Perkinsus chesapeaki, qui infecte les palourdes et les coques.
Ce nouveau mollusque non indigène, envahissant ou non pourrait avoir un impact écologique et/ou économique négatif.
Dans la mer des Wadden (Allemagne) et aux Pays-Bas, M. lateralis semble avoir trouvé les conditions parfaites pour devenir une véritable espèce envahissante et pour supprimer à terme le développement d'autres habitants du sable, comme les mollusques de la mer du Nord Abra alba, Cerastoderma edule, Macoma balthica, Mactra stultorum, Spisula solida et surtout Spisula subtruncata.
Son introduction en Allemagne, aux Pays-Bas et en Belgique est attribuée aux eaux de ballast (Kerckhof, 2019).
Cependant, il est peu probable que cette espèce s'établisse à moins que l'habitat ne soit déjà perturbé, et les impacts peuvent être de courte durée car cette espèce est sujette à des effondrements de population.
Un avantage positif possible est que de nombreuses espèces indigènes pourraient potentiellement utiliser M. lateralis comme ressource alimentaire. Des auteurs, Craeymeersch et al. (2019), prévoient que de nombreuses espèces indigènes de la mer du Nord pourront se nourrir de M. lateralis, notamment des oiseaux de rivage (tels que l'huîtrier-pie (Haematopus ostralegus) , le bécasseau sanderling (Calidris alba) et le bécasseau maubèche (Calidris canutus) et des canards de mer (tels que la macreuse noire (Melanitta nigra), l'eider à duvet (Somateria mollissima) et le fuligule morillon (Aythya fuligula).
En 1968, un auteur (A. Calabrese) a comparé Mulinia lateralis à la drosophile (insecte diptère) des généticiens du fait de sa petite taille, sa fécondité, son développement rapide, de la transparence de sa coquille au début de son développement, de sa courte période de maturation sexuelle et de sa facilité d’élevage au laboratoire. En effet de nombreux laboratoires de génétique exploitent ces capacités pour des expériences de transgenèse. Ils ont ainsi montré que cette espèce présentait une très grande diversité génétique au cours des générations.
Mulinia lateralis est présente du Panama au Maryland (Etats-Unis) dans les roches du Miocène supérieur (- 11 millions d’années) au Pléistocène supérieur (- 11 700 ans).
Mactre naine, cette espèce appartient à la famille des Mactridés et elle est de petite taille.
Mulinia: probablement du grec [mùlinos] = moulin. Le zoologiste britannique John Edward Gray (1800-1875) ne précise pas l’origine de ce nom de genre. Serait-ce un nom propre et donc une dédicace ?
lateralis: du latin [lateralis] = qui tient au côté, des côtés donc probablement pour la crête radiale postérieure. Ce nom a été donné par le zoologiste Thomas Say (1787-1834). qui vivait à Philadelphie (Pennsylvanie, côte est des États-Unis d’Amérique).
Numéro d'entrée WoRMS : 156870
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Bivalvia / Lamellibranchia / Pelecypoda | Bivalves / Lamellibranches / Pélécypodes | Mollusques aquatiques, filtreurs, au corps comprimé latéralement. Coquille composée de 2 valves articulées disposées de part et d’autre du plan de symétrie. Absence de tête, de pharynx, de radula et de glande salivaire. |
| Sous-classe | Autobranchia | Autobranches | |
| Infra-classe | Heteroconchia | Hétéroconchie | |
| Subter-classe | Euheterodonta | Euhétérodonte | |
| Super ordre | Imparidentia | Imparidenties | |
| Ordre | Venerida | Vénérides | Coquille mince, allongée, dure et costulée, bâillante à une ou aux deux extrémités. Ligament à la fois interne et externe. |
| Super-famille | Mactroidea | ||
| Famille | Mactridae | Mactridés | Coquille équivalve, mince, porcelanée. Charnière à dents principales en forme de V. Ligament externe avec resilium, logé dans un chondrophore. Siphons entièrement ou partiellement unis. Sinus palléal arrondi, profond. Pas de byssus. |
| Sous-famille | Mactrinae | Mactrinés | |
| Genre | Mulinia | ||
| Espèce | lateralis |


Vue latérale gauche
La crête radiale postérieure est à droite. L’intervalle entre les deux traits noirs mesure 1 cm.
Individu récolté à Noirmoutier le 14 septembre 2023, sur les plages au sud-ouest de l'île de Noirmoutier, communes de Barbâtre et de la Guérinière par Bruno Boulainghien.
28/02/2025


Vue latérale droite
La crête radiale postérieure est à gauche.
L’intervalle entre les deux traits noirs mesure 1 cm.
Individu récolté à Noirmoutier le 14 septembre 2023, sur les plages au sud-ouest de l'île de Noirmoutier, communes de Barbâtre et de la Guérinière par Bruno Boulainghien.
29/02/2025


Vue des sommets
Coquille globuleuse, les deux crochets sont séparés par un espace.
L’intervalle entre les deux traits noirs mesure 1 cm.
Individu récolté à Noirmoutier le 14 septembre 2023, sur les plages au sud-ouest de l’île de Noirmoutier, communes de Barbâtre et de la Guérinière par Bruno Boulainghien.
28/02/2025


Comparaison des sommets
A gauche : Mulinia lateralis, les deux sommets sont écartés, la coquille est globuleuse.
A droite : Spisula subtruncata, les deux sommets sont très proches.
Les deux photographies ne sont pas au même grossissement. La coquille de droite mesure 2 cm de longueur.
L'individu de gauche a été récolté à Noirmoutier le 14 septembre 2023, sur les plages au sud-ouest de l’île de Noirmoutier, communes de Barbâtre et de la Guérinière par Bruno Boulainghien.
L'individu de droite provient de la plage de Leffrinckoucke (59)
Les deux photos ne sont pas au même grossissement. La coquille de droite mesure 2 cm de longueur.
Macrophotographies (en focus stacking)
28/02/2025

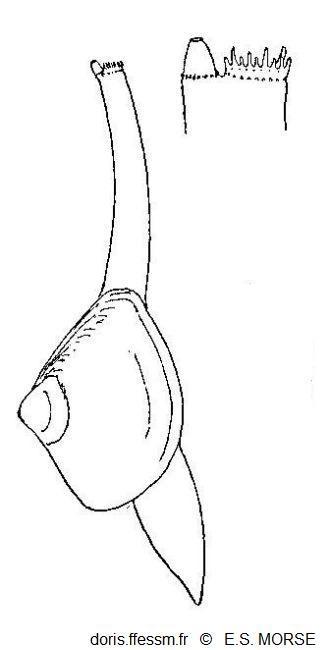
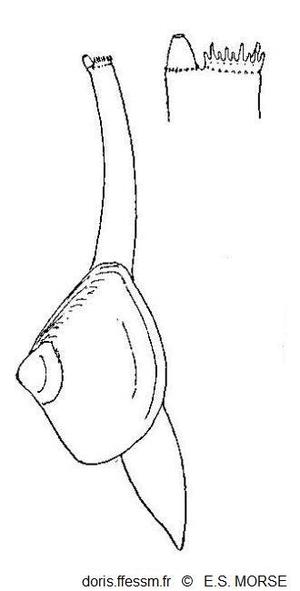
Mulinia lateralis en extension
Le pied et les siphons sont étendus. Les siphons affleurent la surface du sédiment.
Le siphon inhalant porte des papilles et le siphon exhalant montre une valvule conique.
Fig 42 extraite de Morse 1919
Reproduction de documents anciens
1919


Comparaison des caractéristiques de Mulinia lateralis et de Spisula subtruncata
Tableau d'après les caractéristiques des deux espèces
28/02/2025
Rédacteur principal : Yves MÜLLER
Vérificateur : Philippe LE GRANCHÉ
Responsable régional : Yves MÜLLER
Ameye T., 2015, Eerste waarneming van de Amerikaanse strandschelp Mulinia lateralis (Say, 1822) in de Guérande (Frankrijk), De Strandvlo,45(1), 4-10.
Calabrese A., 1968, Mulinia lateralis molluscan fruit fly ?, Proceedings National Shellfisheries Association, 59, 65-66.
Calabrese A.,1970, Reproductive cycle of the coot clam, Mulinia lateralis (Say), in Long Island Sound, The Veliger, 12(3), 265-269.
Chalermwat K., Jacobsen T. R., Lutz R. A., 1991, Assimilation of bacteria by the dwarf surf clam Mulinia lateralis (Bivalvia: Mactridae), Marine ecology progress series. Oldendorf, 71(1), 27-35.
Craeymeersch J. A., Faasse M. A., Gheerardyn H., Troost K., Nijland R., Engelberts A., Perdon K. J., van den Ende D., van Zwol J,. 2019, First records of the dwarf surf clam Mulinia lateralis (Say, 1822) in Europe, Marine Biodiversity Records, 12(1), 5.
Gismann L., Wenke L.K., Uhlir C., Arbizu P.M., Wehrmann A., 2023, Status and occurrence of the non-indigenous dwarf surf clam Mulinia lateralis (Say, 1822) in the central Wadden Sea (southern North Sea)—a systematic survey, Marine Biodiversity, 53, 83, 11p.
Grassle J.P., Snelgrove P.V., Butman C.A.n., 1992, Larval habitat choice in still water and flume flows by the opportunistic bivalve Mulinia lateralis, Netherlands Journal of Sea Research, 3, 33-44.
Guo X., Allen Jr. S. K., 1994, Sex determination and polyploid gigantism in the dwarf surf clam (Mulinia lateralis Say), Genetics, 138(4), 1199-1206.
Holmes A.M., Jarvis S., Delduca E., 2023, First Record for the UK of the non-native Dwarf Surf Clam - Mulinia lateralis (Say, 1822). Journal of Conchology, 44 (6), 575-578.
Huber M., Pelletier M.C., Charles J.B., Burgess R.M., 1992, Ammonia tolerance of the bivalve Mulinia lateralis sublethal sediment toxicity test, Bulletin of Environmental Contamination and Toxicology, 59(2), 292-297.
Kerckhof F., 2019, Mulinia lateralis (Say, 1822) de Kleine Amerikaanse strandschelp nu ook in Belgie, De Strandvlo, 39(1), 4-9.
Kennedy V. S., Mihursky J. A., 1971, Upper temperature tolerances of some estuarine bivalves, Chesapeake Science, 12(4), 193-204.
Klunder L., Lavaleye M., Kleine Schaars L., Dekker R., Holthuijsen S., van der Veer H.W., 2019, Distribution of the dwarf surf clam Mulinia lateralis (Say, 1822) in the Wadden Sea after first introduction, BioInvasions Records, 8(4), 818–827.
Latry L., Jourde J., Bachelet G., Blanchet H., Bocher P., Bujan V., Chenu A., Donnez M., Fouet M., Gouillieux B., Lavesque N., Zaragosi L., Massé C., 2025, First and disturbing records of the bivalve molluscs Potamocorbula amurensis (Schrenck, 1861) and Mulinia lateralis (Say, 1822) in the Bay of Biscay (France), An aod - les cahiers naturalistes de l’Observatoire marin, 15p.
Lu J. K., Chen T. T., Allen S. K., Matsubara T., Burns J. C., 1996, Production of transgenic dwarf surfclams, Mulinia lateralis, with pantropic retroviral vectors, Proceedings of the National Academy of Sciences, 93(8), 3482-3486.
Mann R., Campos B. M., Luckenbach M. W., 1991, Swimming rate and responses of larvae of three mactrid bivalves to salinity discontinuities, Marine Ecology Progress Series, 257-269.
Morse E.S., 1919, Observations on living Lamellibranchs of New England, Proceedings of the Boston Society of Natural History, 35, 139-196.
Nolf F., Mulinia lateralis (Say, 1822) (Mollusca: Bivalvia: Mactridae): comparison between native and introduced populations, and differences with Spisula subtruncata (da Costa, 1788), Présentation Powerpoint.
Say T., 1822, An account of some of the marine shells of the United States, Journal of the Academy of Natural Sciences, Philadelphia. 2(1), 221-248; 2(2), 257-276, 302-325.
Valdez E., Araiza D. N., 2015, Size-, side- and site-related predation of Naticid gastropods on dwarf surf clams (Mulinia lateralis) and incongruous ark clams (Anadara brasiliana) on the Atlantic Coast, DePaul Discoveries, 4(1), 15.
Wells-Berlin A. M., Perry M. C., Kohn R. A., Paynter Jr, K. T., Ottinger M. A., 2015, Composition, shell strength, and metabolizable energy of Mulinia lateralis and Ischadium recurvum as food for wintering Surf Scoters (Melanitta perspicillata), PloS one, 10(5), e0119839.
Yang Z., Zhao A., Teng M., Li M., Wang H., Wang X., Liu Z., Zeng Q., Hu L., Hu J., Bao Z., Huan X., 2024, Signatures of selection in Mulinia lateralis underpinning its rapid adaptation to laboratory conditions, Evolutionary Applications, 17,e13657.
La page de Mulinia lateralis n’existe
pas encore dans l'Inventaire National du Patrimoine Naturel.
La page de Mulinia lateralis sur le site de référence de DORIS pour les mollusques :MolluscaBase
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.







")

