

Animal d’une dizaine de millimètres de longueur
Corps ovale, bombé dorsalement et aplati ventralement
Premier segment thoracique fusionné avec la tête
Péréiopodes (pattes marcheuses) tous semblables sauf les premiers qui se sont transformés en maxillipèdes (appendices buccaux)
Peut se rouler en boule à la manière des cloportes
Uropodes externes présentant des indentations nettes
Au microscope : les péréiopodes et les maxillipèdes sont dotés de soies plumeuses
Sphérome, sphérome dentelé (in Loyer, 1995 : 78)
Toothed sphaeroma (GB)
Oniscus conglobator Pallas, 1766
Sphaeroma conglobator (Pallas, 1766)
Oniscus globator Pallas, 1772
Oniscus serratus J. C. Fabricius, 1787
Manche-mer du Nord, Atlantique, Méditerranée
Zones DORIS : ● Europe (côtes françaises)Originaire des côtes atlantiques et méditerranéennes, l’espèce a progressivement colonisé une grande partie des côtes mondiales. En Atlantique Nord-Est, elle est observée depuis les Açores jusqu’en Irlande.
Elle est rare ailleurs ; elle a été signalée sur les côtes de l’Atlantique Nord-Ouest, de l'Afrique Est et Ouest et de l’océan Indien.
La limite de répartition septentrionale correspond à l’isotherme 9 °C en avril.
L’espèce a été introduite récemment sur les côtes sud-américaines (Brésil, Argentine, Uruguay).
Sphaeroma serratum est une espèce qui vit dans la zone de balancement des marées (espèce intertidale*). On la rencontre sous les pierres ou dans les crevasses des rochers sur l’estran, aux environs de la mi-marée et un peu au-dessus (son optimum se situant juste au-dessous du niveau haut des marées de mortes eaux). Elle fréquente également les massifs de Sabellaria alveolata et les coquilles vides qui sont dans les laisses de mer.
En Méditerranée, elle est une espèce caractéristique de l’habitat EUNIS* A2.13 (biocénoses méditerranéennes du détritique médiolittoral).
En Atlantique, en Manche et mer du Nord, elle est une espèce indicatrice de l’habitat 1140-6 (sédiments hétérogènes envasés). L’espèce se rencontre habituellement en milieu marin pur mais elle ne dédaigne pas les eaux saumâtres comme les estuaires (l’adulte a, expérimentalement, une plage moyenne de tolérance de salinité allant de 2 g/L. à 75 g/L, ce qui en fait clairement une espèce euryhaline*).
Cependant, en milieu naturel, lorsque la salinité baisse, elle est remplacée par d’autres espèces du genre Sphaeroma (comme Sphaeroma hookeri ou Sphaeroma rugicauda) ou Lekanesphaera (autrefois rattaché au genre Sphaeroma).
L'animal adulte mesure une dizaine de millimètres de long rappelant l’allure d’un cloporte. La couleur, qui est souvent unie, varie du beige très clair, presque blanc, au gris foncé.
Le corps, ovale, est bombé dorsalement et aplati ventralement.
Le premier segment thoracique est fusionné avec la tête.
La « queue » (pléotelson*) est arrondie. Les uropodes*, biramés, sont fixés latéralement, les exopodites* (ou endopode) et endopodites (ou exopode) ayant approximativement la même longueur. Les endopodes des uropodes sont arrondis à l’apex*. Les exopodes des uropodes sont mobiles et présentent des indentations très nettes.
Les péréiopodes* sont tous semblables (sauf les premiers, qui se sont transformés en maxillipèdes*) et portent des soies* plumeuses.
Les maxillipèdes ne présentent pas de lobes et portent également des soies plumeuses.
Les antennes sont constituées d’un pédoncule de 5 articles et d’un flagelle* de 15 à 18 articles. Tous les articles du flagelle portent une soie simple à l’extrémité distale (la plus éloignée).
Tous les représentants de la famille des Sphaeromatidés se ressemblent énormément et leur détermination fiable reste complexe. La présence d’indentations sur les exopodes* des uropodes*, observables à l’œil nu, réduit déjà le nombre d’espèces possibles, la seule confusion possible dans le biotope habituel de l’espèce étant Lekanesphaera levii.
La séparation entre les deux espèces nécessite l’utilisation du microscope et est assez immédiate, les soies des péréiopodes de Lekanesphaera levii étant simples alors que celles de Sphaeroma serratum sont plumeuses.
L’observation des maxillipèdes, sans bosse, confirmera la diagnose.
Sphaeroma serratum est, comme tous les représentants de la famille des Sphaeromatidés, un détritivore*, se nourrissant des débris végétaux présents dans sa zone de vie (algues et phanérogames* marines). Des études (Prato & al. 2012) ont montré que cette espèce se nourrissait également de débris animaux, en quantité non négligeable, et de diatomées* lorsque celles-ci prospèrent (donc plutôt lorsque les eaux sont chaudes).
Les sexes sont séparés et il existe un dimorphisme* sexuel. Les mâles sont légèrement plus grands que les femelles et possèdent des antennes plus fortes. Les femelles présentent une allure générale plus globuleuse. Des cas résiduels d’hermaphrodisme* sont cependant signalés dans la littérature.
La fécondation* est interne, les mâles étant dotés de deux stylets copulateurs.
Les œufs sont conservés jusqu’à éclosion dans une poche marsupiale* présente
sur l’abdomen de la femelle, formée par des oostégites*.
Sur les côtes du Finistère, les premières éclosions ont lieu au mois de juin et
les dernières en septembre. La période effective de reproduction est plus
étendue dans les eaux chaudes, où les femelles se reproduisent une fois l’année
de leur naissance puis deux fois l’année suivante. Les jeunes larves* débutent
leur croissance au sein de la poche marsupiale, qu’elles quittent lorsqu’elles
atteignent une taille d’environ 2 mm. A ce stade, les larves ressemblent déjà à
l’adulte mais ne possèdent que 6 paires de pattes thoraciques (contre 7 chez
les adultes).
La poche marsupiale joue notamment un rôle de protection contre
les variations de salinité du milieu ambiant, les jeunes larves ne possédant
pas de capacités d’osmorégulation (régulation des concentrations en sels dissous dans les liquides de l'organisme).
Les jeunes individus sont sexuellement indifférenciés. Il y a plusieurs mues*
de croissance avant une mue de différenciation sexuelle, qui a lieu entre 2
mois et 4 mois après avoir quitté leur mère, ce délai dépendant de la
température de l’eau.
Les mâles adultes ne muent plus après la mue de puberté, après laquelle ils
vivront encore de six mois à un an. Les femelles peuvent vivre plus longtemps
que les mâles.
Sphaeroma serratum est fréquemment rencontré avec un petit isopode commensal, Jaera (Metajaera) hopeana beaucoup plus petit, que l’on rencontrera sur la face inférieure des Sphaeroma, habituellement entre les péréiopodes*.
Sphaeroma serratum est régulièrement infesté par un protozoaire, Palavascia sphaeromae. Ce protozoaire se développe dans le proctodeum (partie terminale du système digestif) de son hôte. Les animaux sains se contaminent en mangeant de la nourriture contaminée par un autre animal. La reproduction du protozoaire est généralement synchronisée avec la mue de l’hôte, même si ceci n’est pas absolument indispensable compte tenu de la localisation du parasite à l’extrémité du système digestif. La relation entre le protozoaire et le crustacé isopode s’apparente davantage à du commensalisme* qu’à du parasitisme*. Le protozoaire se fixe sur les parois internes du proctodeum sans les pénétrer. Le taux de contamination peut être très élevé, puisqu’il atteint 95 % certains mois dans l’étang de Thau.
Sept phénotypes* ont été identifiés en Bretagne chez cette espèce par Bocquet, Lévi et Tessier en 1951 (voir bibliographie), conditionnés par 4 couples d’allèles (version variable d'un même gène). Cinq d’entre eux sont structuraux :
Ces types ne sont pas intégralement retrouvés dans les populations en limite de répartition septentrionale, ni dans les populations correspondant à des zones d’introduction anthropique* de l’espèce. Ces phénotypes très marqués ont permis la réalisation de nombreuses études sur les populations de cette espèce. Les populations sont caractérisées par des pourcentages propres de chaque phénotype, qui restent constants d’une génération à l’autre mais peuvent varier fortement sur une faible distance géographique (cas des populations rencontrées sur les côtes finistériennes). Les études réalisées ont par ailleurs montré qu’il y avait une réelle panmixie* chez cette espèce (les couples ne se forment pas en fonction de leur phénotype et tous les allèles sont compatibles).
La dessalure des eaux de l’étang de Berre dans les années 1970, liée à la mise en service de l’usine hydroélectrique de Saint-Chamas, a provoqué la disparition de Sphaeroma serratum de cet étang, espèce à laquelle s’est substituée Lekanesphaera hookeri, espèce plus euryhaline que la précédente. Cette espèce a elle-même fini par fortement régresser dans l’étang compte tenu de la poursuite des apports d’eau douce et de l’apparition de zones anoxiques*. La modification de l’exploitation de l’usine hydroélectrique engagée au milieu des années 2000 a permis d’améliorer la situation.
Sphaeroma serratum est une espèce relativement résistante à la pollution, nettement plus que Idotea balthica par exemple, et plusieurs études ont été réalisées pour utiliser cet isopode comme bio-indicateur de pollution aiguë aux métaux lourds. Ces études ont donné des résultats probants, mais il n’existe pas actuellement de protocole normalisé fondé sur ce crustacé.
Régina Wetzer & al. ont démontré récemment (2018) que la famille des Sphaeromatidés n’est pas monophylétique*. Cependant, au sein de cette famille, l’étude a montré que le genre Sphaeroma était monophylétique et constituait un clade frère de Lekanesphaera. Cette étude conforte donc, parmi toutes ses conclusions, la validité des genres Sphaeroma et Lekanesphaera.
Sphérome denté : simple traduction du nom scientifique.
Le nom du genre, Sphaeroma, provient du grec ancien [sphairoma] = quelque chose d’arrondi, allusion directe à la faculté de cette espèce de se mettre en boule en cas de danger.
Le nom d’espèce serratum provient du latin [serratus] = dentelé. Ce nom a été donné par l'entomologiste danois Johan Christian Fabricius (1745-1808) en 1787 pour souligner la présence d’indentations sur les exopodes* des uropodes*.
Numéro d'entrée WoRMS : 118973
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Arthropoda | Arthropodes | Animaux invertébrés au corps segmenté, articulé, pourvu d’appendices articulés, et couvert d’une cuticule rigide constituant leur exosquelette. |
| Sous-embranchement | Crustacea | Crustacés | Arthropodes à exosquelette chitineux, souvent imprégné de carbonate de calcium, ayant deux paires d'antennes. |
| Super classe | Multicrustacea | ||
| Classe | Malacostraca | Malacostracés | 8 segments thoraciques, 6 segments abdominaux. Appendices présents sur le thorax et l’abdomen. |
| Sous-classe | Eumalacostraca | Eumalacostracés | Présence d’une carapace recouvrant la tête et tout ou partie du thorax. |
| Super ordre | Peracarida | Péracarides | Les femelles sont dotées d'une cavité d'incubation formée par des expansions lamelleuses des péréiopodes. |
| Ordre | Isopoda | Isopodes | Corps comprimé dorso-ventralement, première paire d’antennes beaucoup plus petite que la seconde, yeux non pédonculés. 7 paires de pattes de même apparence. |
| Sous-ordre | Sphaeromatidea | Sphaeromatidés | |
| Famille | Sphaeromatidae | Sphaeromatidés | |
| Genre | Sphaeroma | ||
| Espèce | serratum |
") Crustacés Malacostracés (crabes, crevettes...)
Crustacés Malacostracés (crabes, crevettes...)

Quelques individus
Vue d’ensemble de quelques individus trouvés sous une pierre. Les indentations sur les exopodes des uropodes sont visibles sur le plus gros individu sombre.
Sur l'estran, Macrophoto, rapport 1:1, Pointe de la Rognouse, Binic (22)
16/07/2022
Crustacés Malacostracés (crabes, crevettes...)

Quelques individus
Cette espèce peut présenter des variations de couleur entre individus très importantes.
Sur l'estran, Macrophoto, rapport 1:1, Pointe de la Rognouse, Binic (22)
16/07/2022


Phénotype Lunulatum
Un individu présentant le phénotype Lunulatum, avec des taches blanches frontales, latérales et caudales. Ce phénotype est porté par 6 génotypes possibles.
Sur l'estran, Macrophoto, rapport 1:1,
Pointe de la Rognouse, Binic (22)
16/07/2022


Phénotype Discretum
Un individu présentant le phénotype Discretum, tacheté. Ce phénotype est porté par 2 génotypes.
Sur l'estran, Macrophoto, rapport 1:1,
Pointe de la Rognouse, Binic (22)
16/07/2022


Phénotype Rubrum
Sur cette photographie, on aperçoit un
individu arborant une coloration rouge (Rubrum).
Sur l'estran, Macrophoto, rapport 1:1, Pointe de la Rognouse, Binic (22)
16/07/2022


La vie en communauté
Sur ce pare-battage constamment immergé les sphéromes partagent leur habitat avec de nombreux autres organismes (ascidies coloniales, bryozoaires, etc.).
port de Hyères (83)
09/12/2022


Plétotelson et uropodes
On distingue bien ici le pléotelson plutôt lisse et uniforme ainsi que les uropodes donc les exopodes ont des marges fortement dentées.
Port de Hyères (83)
09/12/2022


Uropodes
Vue au microscope des uropodes d’un individu. On voit très bien les indentations marquées sur l’exopode et l’apex arrondi de l’endopode.
Photo prise au microscope à partir d’un individu prélevé sur l’estran à la Pointe de la Rognouse, Binic (22)
16/07/2022

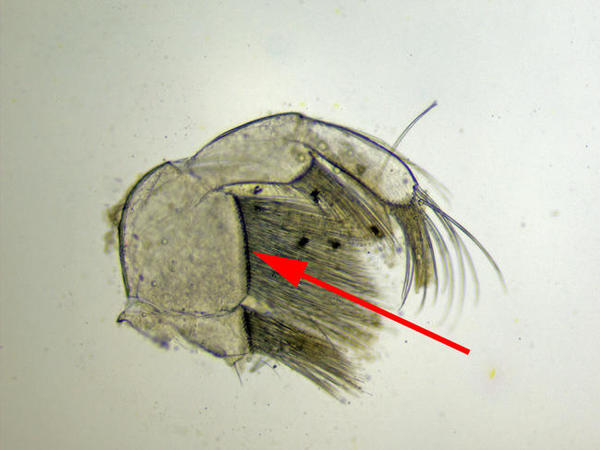
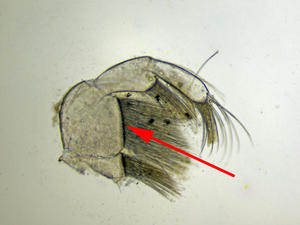
Maxillipède
Vue au microscope d’un maxillipède. La flèche montre le segment 2 dépourvu de bosse et orné de nombreuses soies
Photo prise au microscope à partir d’un individu prélevé sur l’estran à la Pointe de la Rognouse, Binic (22)
17/07/2022

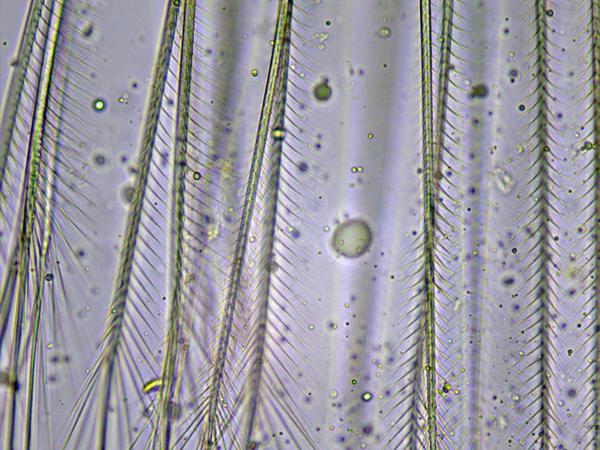
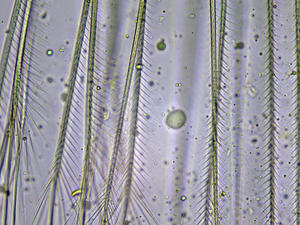
Soies plumeuses
Chez Sphaeroma serratum, les soies des maxillipèdes et des péréiopodes sont plumeuses comme on peut le voir ici.
Photo prise au microscope à partir d’un individu prélevé sur l’estran à la Pointe de la Rognouse, Binic (22)
17/07/2022

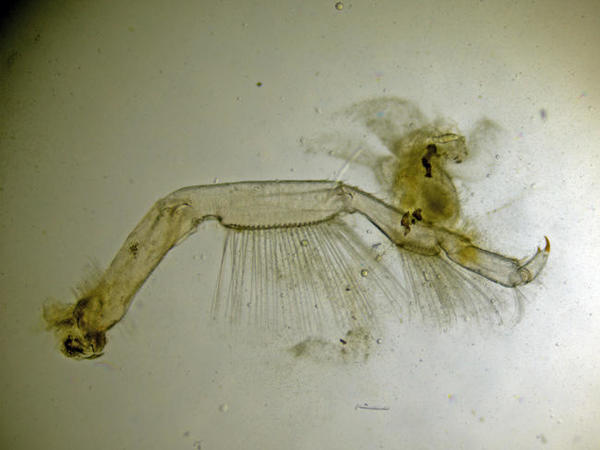
Un péréiopode
Vue au microscope d’un péréiopode. Les soies sont plumeuses (non visible à ce grossissement).
Photo prise au microscope à partir d’un individu prélevé sur l’estran à la Pointe de la Rognouse, Binic (22)
17/07/2022


Volvation
Lorsque l’animal est importuné, il se roule en boule, n’offrant qu’une prise réduite à l’intrus et bloquant l’accès aux parties molles. Ce comportement s’appelle la volvation. Les yeux, noirs, à facettes sont bien visibles.
Macrophoto, rapport 1:1 agrandie, Binic (22)
25/07/2022


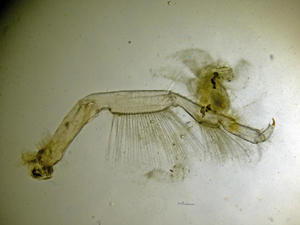
Vue de dessous
Vue de dessous d’un animal montrant les paires de pattes marcheuses ou péréiopodes.
Sur l'estran, macrophoto, rapport 1:1, Pointe de la Rognouse, Binic (22)
17/07/2022


Le biotope
Vue d’ensemble du biotope typique où cette espèce pourra être rencontrée, en grand nombre, en Manche/Atlantique (il s’agit de l’habitat 1140-6 (sédiments hétérogènes envasés). Dans la partie haute de la photo, proche ou au-dessus du niveau haut des marées, les sédiments ne sont pas assez humides pour héberger cette espèce (on est alors dans l’habitat 1140-2- galets et cailloutis des hauts de plage à Orchestia).
L'estran, Binic, Pointe de la Rognouse (22)
17/07/2022


Vue de dos
Sur un pilier en bois d'un ponton
Port-Mahon-Etang, Sigean-la-Berre, 0,2 m
02/12/2022


Vue de côté
Sur un pilier en bois d'un ponton
Port-Mahon-Etang, Sigean-la-Berre, 0,2 m
02/12/2022
Rédacteur principal : Christophe QUINTIN
Vérificateur : Pierre NOËL
Responsable régional : Yves MÜLLER
Bocquet C., Lévi C., Teissier G., 1951, Recherches sur le polychromatisme de « Sphaeroma serratum » F, Archives de Zoologie Expérimentale, 87, 245-297.
Bocquet C., Teissier G., 1960, Génétique des populations de Sphaeroma serratum (F.), I, Stabilité du polychromatisme local, Cahiers de biologie marine, 1(1), 103-111.
Bocquet C., Lejuez R., Teissier G., 1960, Génétique des populations de Sphaeroma serratum (F.), III, Comparaison des populations mères et des populations filles pour les Sphéromes du Cotentin, Cahier de biologie marine, 1, 279-294.
Bocquet C., Lejuez R., Teissier G., 1965, Génétique des populations de Sphaeroma serratum (F.), VI, Mise en évidence de la panmixie chez Sphaeroma serratum, Cahier de biologie marine, 6, 195-200.
Bocquet C., Lejuez R., Teissier G., 1966, Génétique des populations de Sphaeroma serratum (F.). VII Données complémentaires sur la panmixie, Cahier de biologie marine, 7, 23-30.
Cafaro M.J., 2000, Gut fungi of isopods : the genus Palavascia, Mycologia 92(2), 361-369.
Charmantier G., 1974, Le cycle biologique de Sphaeroma serratum (Fabricius, 1787) (Crustacea, Isopoda, Flabellifera) dans l'étang de Thau (Golfe de Lion), Vie et Milieu, 24, 285-300.
Charmantier G., Manier J.F., 1981, Relations écologiques entre Sphaeroma serratum (Fabricius, 1787) (Crustacea, Isopoda, Flabellifera) et son commensal intestinal Palavascia sphaeromae Tuzet et Manier, 1948, ex Manier, 1968 (Trichomycètes, Eccrinales, Palavasciaceae), Vie et Milieu, 31, 101-111.
Charmantier G., Charmantier-Daures M., 1994, Ontogeny of osmoregulation and salinity tolerance in the isopod crustacean Sphaeroma serratum, Marine Ecology Progress Series, 114, 93-102.
El-Din M.IS., Sakiko Y., Mohamed S.Z., Bedir M.A., Bahgat I.M., Nishimura O., 2014, Investigating the use of Sphaeroma serratum (Crustacea, Isopoda) as bio-indicator for heavy metal pollution in Lake Timsah, Suez Canal using alkaline comet assay technique, Egyptian Academic Journal of Biological Sciences, 6(1), 7-26.
Fabricius J. C., 1787, Mantissa Insectorum : Sistens Eorum Species Nuper Detectas, Adjectis Characteribus Genericis, Differentiis Specificis, Emendationibus, Observationibus, Tome I. Hafniae, Christ. Gottl. Proft., page 242.
Hoestlandt H., 1958, Comparaison des fréquences raciales d’un crustacé littoral, Sphaeroma serratum, aux Canaries et sur d’autres côtes atlantiques insulaires ou continentales, Anuario de estudios atlanticos, 4, 1-20.
Holthuis L.B., 1950, Isopodes et Tanaidacés marins de la Belgique ; remarques sur quelques espèces de la zone méridionale de la Mer du Nord, Bulletin de l’Institut royal des Sciences naturelles de Belgique, Tome XXVI, 53, 1-19.
Jacobs B.J.M., 1987, A Taxonomic Revision of the European, Mediterranean and NW African species generally placed in Sphaeroma Bosc, 1802. Isopoda : Flabellifera : Sphaeromatidae, Zoologische Verhandelingen Leiden, 238, 1-71.
Kerambrun P., Szekielda K.H., 1969, Influence des modifications hydrologiques sur le peuplement de Sphaeroma dans l’étang de Berre, Cahiers de biologie marine, 10, 301-308.
Kerambrun P., 1970, Remplacement de Sphaeroma serratum par S. hookeri dans l'Etang de Berre par suite de sa dessalure, Marine Biology, 6, 128–134.
Manier J.F., 1979, Etude ultrastructurale de Palavascia sphaeromae parasite du proctodeum de Sphaeroma serratum, Annales de parasitologie, 54(5), 537-554.
Prato E., Biandolino F., Scardicchio C., 2006, Test for acute toxicity of Copper, Cadmium and Mercury in five marine species, Turkish Journal of Zoology, 30, 285-290.
Prato E., Danieli A ., Maffia M., Biandolino F., 2012, Lipid Contents and Fatty Acid Composition of Idotea baltica and Sphaeroma serratum (Crustacea : Isopoda) as Indicators of Food Sources, Zoological Studies, 51(1), 38-50.
Rezig M., 1979, Etude comparée du cycle biologique de quelques espèces du genre Sphaeroma (Isopodes flabellifères) en Tunisie, Bulletin de l’Institut National scientifique et technique d’océanographie et de pêche de Salammbô, 6(1-4), 93-121.
Schlund E., Basuyaux O., Lecornu B., Pezy J.P., Baffreau A., Dauvin J.C., 2016, Macrofauna associated with temporary Sabellaria alveolata reefs on the west coast of Cotentin (France), SpringerPlus 5, 1260.
Teissier G., 1960, Génétique des populations de Sphaeroma serratum (F.). II Calcul des fréquences géniques, Cahiers de biologie marine, 1, 221-230.
Wetzer R., Bruce N.L., Pérez-Losada M., 2018, Relationships of the Sphaeromatidae genera (Peracarida: Isopoda) inferred from 18S rDNA and 16S rDNA genes, Arthropods systematics and phylogeny, 76(1), 1-30.La page de Sphaeroma serratum dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.















")

