

Grand calmar 80 cm, voire 100 cm, de longueur du manteau
Manteau cylindrique fusiforme, épais
Nageoires en forme de losange sur toute la longueur du manteau
Tête, aussi large que le manteau, avec 2 gros yeux
8 bras et 2 tentacules
Bras courts avec 2 rangées de ventouses
pédonculées
Massue tentaculaire à 4 rangées de ventouses
Présence d’une membrane sur les 3 paires de bras supérieurs
Couleur rouge, rose, brun clair ou jaune clair
Chipiloua
Diamond squid, diamondblack squid, rhomboid squid (GB), Chipiron, chipiron volantin (E), Rhomben-Kalmar (D), Totano rombo
(I)
Comme cette espèce est présente
dans de vastes régions, elle porte de nombreux noms dans les pays qui l’exploitent,
comme le Japon (sode-ika, taru-ika, hako-ika, kasa-ika, ushi-ika, sei-ika,...).
Sepioteuthis major Gray, 1828
Thysanoteuthis elegans Troschel, 1857
Cirrobranchium filiferum Hoyle, 1904
Thysanoteuthis nuchalis Pfeffer, 1912
Cirrobranchium danae Joubin, 1933
Thysanoteuthis danae (Joubin, 1933)
Cosmopolite des eaux tropicales et sub-tropicales
Zones DORIS : ● Europe (côtes françaises), ○ [Méditerranée française], ○ [Atlantique Nord-Est, Manche et mer du Nord françaises], ● Indo-Pacifique, ● CaraïbesCette espèce est cosmopolite des eaux tropicales et sub-tropicales des océans, de la Méditerranée et de la mer des Caraïbes. Transporté par les courants chauds, ce calmar peut être présent à des latitudes plus élevées.
Thysanoteuthis rhombus est une espèce pélagique* qui est présente dans les eaux océaniques ouvertes de plus de 400 m de profondeur où la température de l'eau de surface est généralement supérieure à 20 °C (23 à 26 °C). Ce calmar vit dans les eaux proches de la surface de 0 à 50 m (épipélagique*) pendant la nuit et les eaux intermédiaires de 400 à 800 m (mésopélagique*) pendant le jour.
Ces calmars se déplacent par 2 (en couples ?) ou en très petits groupes.
Ce calmar est de grande taille, son manteau* mesure généralement 80 cm de longueur. La longueur du manteau est la mesure de longueur utilisée chez ces animaux (la tête et les bras ne sont pas pris en compte dans cette mesure). Certains exemplaires peuvent mesurer 100 cm voire plus. Adulte, il peut peser de 24 à 30 kg.
Le manteau cylindrique fusiforme est légèrement aplati dorso-ventralement (ovale en section transversale) avec une extrémité postérieure arrondie. Ce manteau épais (4-5 cm d’épaisseur chez les grands calmars de 70-80 cm de longueur du manteau) est très musclé (Nigmatullin & al.,1998).
Les nageoires de cette espèce sont caractéristiques : elles forment un losange (ou rhombe) et s’étendent sur toute la longueur du manteau. La plus grande largeur se situe entre les 2/3 et les 3/4 de la longueur des nageoires.
La tête, aussi large que le manteau, porte 2 gros yeux sans membrane cornéenne (caractéristique de l'ordre des Œgopside). Elle porte 8 bras et 2 tentacules* autour de la bouche.
Les bras sont relativement courts et de longueur différente par paires. Ils sont fortement musclés avec 2 rangées de ventouses pédonculées. A la base de chaque ventouse, il y a un prolongement que l'on nomme cirre*, trabécule, frange. Ces cirres sont de taille différente selon la paire de bras. Ils sont reliés par une membrane sur les 3 paires de bras supérieurs (Troschel, 1857).
Chez le mâle adulte, le bras ventral gauche est transformé en bras copulateur (bras hectocotyle*).
Les 2 tentacules portent à leur extrémité, comme chez tous les calmars, une massue avec 4 rangées de ventouses. Ces tentacules ne sont visibles qu'au moment de la capture des proies ou quand l'animal est mort.
La couleur de la peau est rouge, rose, brun clair ou jaune clair. Chez les adultes, il y a souvent une bande longitudinale rouge sur la face dorsale du manteau. Cependant, les T. rhombus nageant sous les lumières de pêche, la nuit, ont des tissus corporels blanchâtres ou semi-transparents (Nigmatullin, 1998).
La forme du corps de cette espèce est proche de celle des Loliginidés (comme Loligo vulgaris) dans ses proportions du manteau, de la tête et des bras. Mais Thysanoteuthis rhombus n'a pas de membrane cornéenne sur l'œil et est proche des Ommastrephidés (ordre des Œgopsides). La forme des nageoires est très caractéristique.
Peu de genres de calmars ont une nageoire aussi longue que le manteau.
Les calmars capturent leur proie en faisant jaillir les deux tentacules munis de ventouses uniquement sur les massues et la ramènent dans la couronne des bras pour la maintenir. Leur bec corné (bec de perroquet) sert à déchiqueter la proie. Leur salive contient des substances neurotoxiques*. La radula* sert à convoyer la nourriture.
Les juvéniles se nourrissent de crustacés (dont des copépodes), de petits céphalopodes et de larves* de poissons dans les eaux proches de la surface.
Les subadultes* et les adultes se nourrissent principalement pendant la journée dans les couches d'eau profondes, surtout entre 400 et 650 m, sur des concentrations denses de petits poissons de profondeur mésopélagiques* peu actifs comme Argyropelecus affinis, Chauliodus sloani, des Myctophidés, etc., des calmars comme Abraliopsis sp., Ctenopteryx sp., Dosidiscus gigas, Todarodes pacificus, d’animaux macroplanctoniques* et micronectoniques* et un peu pendant la nuit dans les couches de surface.
Cette espèce est un prédateur actif opportuniste mais elle ne poursuit jamais ses proies. Les proies sont de petite taille (généralement pas plus de 5 à 10 % de la longueur du calmar). Thysanoteuthis rhombus changerait facilement ses habitudes. Il peut chasser des calmars relativement grands, sexuellement mâtures, lorsqu'ils sont inactifs et des poissons en train de frayer.
Les céphalopodes sont des animaux gonochoriques* stricts, ce qui signifie que les sexes sont séparés.
Les mâles et les femelles atteignent la maturité à l'âge de 5 à 8 mois (soit 25 à 35 cm de longueur du manteau* pour les mâles et 50 à 62 cm pour les femelles) (Nigmatullin & al., 1998).
Chez Thysanoteuthis rhombus, l'appariement des mâles et des femelles se produit à un stade immature (la longueur du manteau est inférieure à 10 cm de longueur). C’est le seul céphalopode connu à former des paires composées d’un mâle et d’une femelle (monogamie) qui durent depuis avant la maturation sexuelle et pendant toute la période de reproduction (Bello, 2006, Nigmatullin & al.,1998).
Les calmars présentent un dimorphisme sexuel visible y compris chez les individus immatures : la troisième paire de bras est deux fois plus longue chez les mâles que chez les femelles.
L'accouplement a lieu dans la position "tête contre tête". Le mâle, avec son bras hectocotyle*, porte ses spermatophores* sur la membrane buccale de la femelle.
La période de reproduction dure toute l'année dans les eaux tropicales et pendant la saison chaude (fin de l’été et au début de l’automne) dans les régions périphériques comme la Méditerranée.
Au moment de la ponte, près de la surface ou en profondeur, les femelles sécrètent un mucus qui gonfle au contact de l’eau. Elles façonnent cette sécrétion en un cylindre. Deux fils muqueux contenant chacun une rangée d’ovules* forment un seul cordon contenant une double rangée d’ovules. Ce cordon sort par l’entonnoir et les spermatozoïdes* stockés sur la membrane buccale assurent la fécondation. Le cordon d’œufs est alors enroulé autour du cylindre. Chaque œuf, de 1,6 à 1,8 mm de diamètre, est contenu dans une capsule transparente de 2,8 à 3,4 mm de diamètre. Une femelle peut produire 8 à 12 masses d’œufs.
Les larves et les juvéniles de cette espèce sont très caractéristiques :
La durée de vie, mesurée à l’aide des statolithes (concrétions calcaires paires situées dans la tête présentant des stries d’accroissement), est d’environ 1 an.
Thysanoteuthis rhombus a un mode de vie unique parmi les calmars. Les mâles et les femelles immatures forment des couples. Ils restent probablement ensemble de façon constante pendant leur vie (monogamie).
Cette espèce peut former de petits bancs, localement communs, de 4 à 28 individus. Des observateurs sous-marins ont précisé que chaque couple était bien séparé des autres.
Les individus de longueur de manteau de 15 à 30 cm sont peu parasités (sauf par des métacercaires* de trématodes). L’infection massive a lieu à partir de 40 cm de longueur du manteau, probablement du fait que Thysanoteuthis rhombus se nourrit de proies relativement petites, contrairement aux autres calmars pélagiques (comme les Ommastrephidés)
Puis ce sont alors des larves de nématodes (Anisakis sp., Porroeaeeum sp.), de cestodes (Nybelinia sp., Tentacularia coryphaenae, Phyllobotrium sp.l., Scolex pleuronectes) et des trématodes didymozoïdes qui parasitent le calmar (Nigmatullin & al., 1985). Certaines espèces de parasites (nématodes, cestodes et trématodes) peuvent être présentes par centaines voire par milliers d’individus par hôte.
Tous ces parasites achèveront leur développement ensuite dans les hôtes finaux qui consommeront Thysanoteuthis rhombus.
Les calmars, comme les seiches, possèdent deux façons de se déplacer.
Lors de leur fuite en arrière, comme les autres calmars, T. rhombus distrait les prédateurs en utilisant un nuage d'encre noir brunâtre.
Les rapides changements de couleurs sont assurés par l’expansion de cellules pigmentées, les chromatophores*. Ces derniers sont petits quand ils sont au repos et grands quand les muscles se sont contractés. Quand les fibres musculaires des chromatophores cessent de se contracter le chromatophore revient à son état (petit) initial.
Il y a un photophore* interne à proximité de l’anus et de la poche à encre chez les spécimens jeunes.
L’appareil de fixation de l’entonnoir au manteau est cartilagineux, robuste et est en forme de T (système de “bouton-pression”).
La rigidité du corps est en partie assurée par une mince lame chitineuse* transparente, la plume (ou gladius). Cette structure correspond à la base chitineuse d'une coquille régressée dont la calcification a été perdue.
Le développement des membranes protectrices et des bras relativement longs au stade paralarvaire peuvent être des adaptations pour flotter dans les courants océaniques plutôt que pour nager activement, et suggèrent que T. rhombus pourrait avoir une longue phase planctonique (Wakabayashi & al., 2005).
Ce grand calmar malgré son apparence et sa puissante musculature est un animal à faible activité et relativement passif pendant la majeure partie de sa vie. Il nage plutôt lentement (jusqu’à 3,5 km/h) en faisant onduler ses nageoires. Cependant, en cas de danger, il est capable d'effectuer une ou plusieurs contractions puissantes de son manteau musculeux et ainsi d'échapper à la situation dangereuse mais pendant une courte durée (Nigmatullin & al., 1995). Certains auteurs sont signalé que cette espèce sautait sur le pont des bateaux. Cette particularité a été contredite sérieusement par d’autres (Nishimura 1966) sauf pour les juvéniles.
Cependant, ce style de vie à faible consommation d’énergie lui permet d’avoir un taux de croissance parmi les plus élevés chez les calmars. A l’âge de 300 jours, il atteint 80 cm de longueur du manteau et plus de 17 kg de poids corporel.
Thysanoteuthis rhombus est un calmar qui est particulier car peu abondant avec une faible densité de population (moins de 5 à 20 kg/km² ou 0,01 à 100 spécimens par km²) et aux déplacements lents. C’est certainement lié au fait que les individus, pour la reproduction, vivent en couples depuis leur stade juvénile jusqu’à leur mort.
Ces stratégies de reproduction et de comportement seraient efficaces pour survivre dans ce milieu particulier.
En raison de sa chair ferme et savoureuse, T. rhombus est ciblé par les pêcheries en expansion dans les régions proches du sud et du centre du Japon. La pêche commerciale de cette espèce se fait au Japon et localement à Taiwan. Sa valeur commerciale élevée en fait une espèce cible importante dans ces eaux. Plusieurs techniques différentes sont employées pour la pêche de cette espèce.
Les îles Canaries seraient également une zone de reproduction pour cette espèce qui pourrait être une espèce cible présentant un intérêt commercial (Pérez & al., 2012). D'autres régions comme la Guadeloupe et la Nouvelle-Calédonie réalisent des études dans le même but.
De nombreux prédateurs se nourrissent de T. rhombus : diverses espèces de calmars Ommastrephidés (pour les jeunes), 2 espèces de coryphènes (Coryphaena equiselis et C. hippurus), des poissons lancettes (du genre Alepisaurus), plusieurs espèces de thons (dont : Thunnus albacares, T. obesus, T. alalunga, Katsuwanus pelamis), l'espadon Xiphias gladius, l’escolier serpent Gempylus serpens, et des requins. Des mammifères marins le consomment également comme les dauphins, dont le dauphin à dents rudes Steno bredanensis, la fausse orque Pseudorca crassidens et le cachalot Physeter macrocephalus.
Les céphalopodes ont tendance à concentrer
rapidement les métaux lourds (ou éléments traces métalliques) et autres
substances toxiques dans leurs tissus, ce qui joue un rôle important dans la
bioaccumulation de ces polluants chez les prédateurs marins et a des
implications pour la consommation humaine. Des niveaux élevés de cadmium et de
mercure sont souvent enregistrés dans les tissus mais uniquement dans la glande digestive (qui ne se consomme pas). Il n’y a aucune accumulation de métaux dans les tissus musculaires (le manteau qui se mange).
Les juvéniles possèdent dans la cavité du manteau un photophore* qui disparaît chez les adultes. Il servirait probablement pour le rapprochement du futur couple.
Ces axones géants ont été observés par L.W. Williams en 1909 et redécouverts par J.Z. Young en 1939. Cette caractéristique particulière a permis l'étude du fonctionnement des messages nerveux par sir A. Huxley et sir A. Hodgkin (tous les deux prix Nobel en 1963 pour ces travaux).
Les calmars sont répartis dans deux ordres : les Myopsides et les Œgopsides, selon la présence (Myopsides) ou l’absence (Œgopsides) d’une membrane cornéenne transparente couvrant l’œil. Les Ommastrephidés sont des Œgopsides.
Aucune information sur l'origine du nom français chiloupa.
Calamar : du latin [calamus] = calame, roseau. La plume (ou gladius) étant comme le roseau taillé pour l'écriture dans l'Antiquité. Ce terme est, de nos jours, davantage utilisé sur les étalages des poissonniers et en gastronomie, calmar serait l'appellation utilisée par les naturalistes.
Thysanoteuthis : du grec [thysanos] = frange, pompon, gland, pour les cirres (ou franges ou trabécules) sur les bras et et du grec [teuthis] = calmar.
rhombus : du grec [rhombos] = rhombe, losange, pour la forme des nageoires.
Numéro d'entrée WoRMS : 141680
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Cephalopoda | Céphalopodes | Yeux complexes, coquille interne, externe cloisonnée ou absente, cavité palléale musclée, siphon musculeux, tentacules ou bras (munis de ventouses). |
| Sous-classe | Coleoidea | Coléoïdes | Ventouses sur les bras, au nombre de 8 au moins. Bras hectocotyle chez le mâle pour le transfert des spermatophores. 2 branchies, 2 néphridies, une coquille interne ou vestigiale, des chromatophores, une poche à encre, un grand cerveau. |
| Ordre | Oegopsida | Oegopsides | Calmars ne possédant pas de membrane cornéenne transparente recouvrant l'œil. Les bras et les tentacules portent des ventouses et/ou des crochets. Cet ordre comprend la plupart des calmars pélagiques et profonds (25 familles). |
| Famille | Thysanoteuthidae | Thysanoteuthidés | Présence à la base des ventouses d'uin petit prolongement (cirre, trabécule) reliés par une membrane sur les 3 paires de bras supérieurs |
| Genre | Thysanoteuthis | ||
| Espèce | rhombus |
 Céphalopodes
Céphalopodes

De gros yeux
La tête, aussi large que le manteau, porte 2 gros yeux sans membrane cornéenne (caractéristique de l'ordre des Œgopside). La nageoire losangique est caractéristique.
Au large de Cannes (06), 30m, de nuit
17/09/2020
Céphalopodes

Posture d'intimidation
Cet individu rencontré au dessus du canyon de Cannes était immobile, en pleine eau, à 30m de profondeur. Passif, il n'a quasiment pas bougé et s'est laissé photographié ! Il a préféré prendre cette posture d'intimidation, tentacules déployés et parfaitement symétriques, que de fuir.
Au large de Cannes (06), 30m, de nuit
17/09/2020


Une forme de losange
Les nageoires de cette espèce sont caractéristiques : elles forment un losange (ou rhombe) et s’étendent sur toute la longueur du manteau. les cirres sur les tentacules sont bien visibles.
Sur cette photo, la bande longitudinale rouge sur la face dorsale du manteau est particulièrement visible.
Au large de Cannes (06), 30m, de nuit
17/09/2020


Des cirres sur les bras
Les bras supérieurs de cette espèce portent des cirres reliés par une membrane.
Au large de Cannes (06), 30m, de nuit
17/09/2020

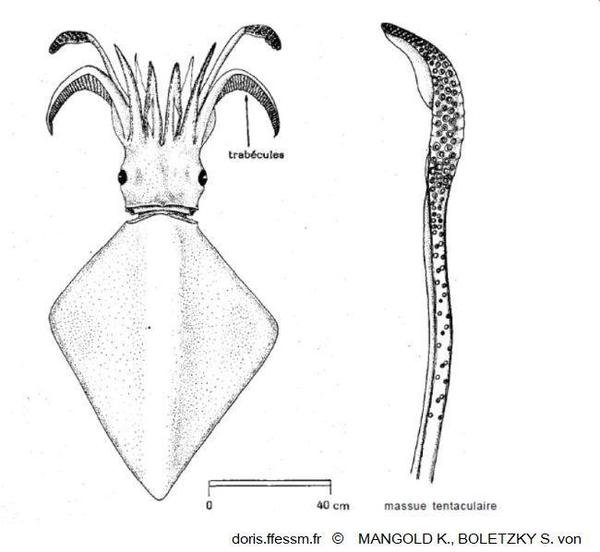
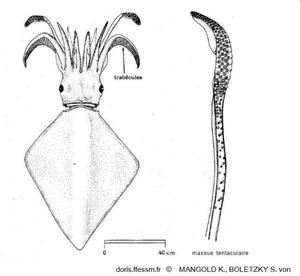
Dessin en vue dorsale de Thysanoteuthis rhombus et d'une massue tentaculaire
Les deux nageoires forment un losange et les cirres (ou trabécules) sur les bras dorsaux sont caractéristiques de cette espèce.
Dessins extraits de Mangold K. et Boletzky S. von, 1987 page 699
Reproduction de documents anciens
1987

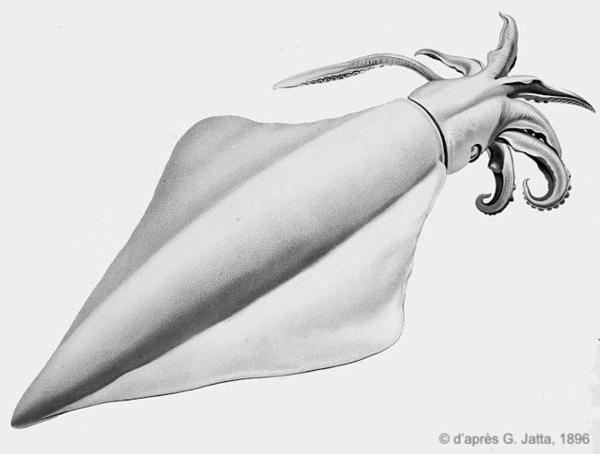
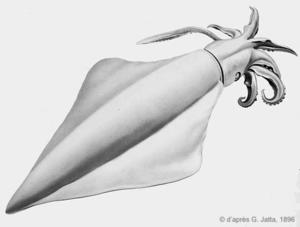
Dessin ancien
Ce dessin montre les grandes nageoires caractéristiques de cette espèce.
Planche N°9. Jatta G., 1896
Reproduction de documents anciens
1896


Ponte de Chipiloua commun près de la surface (10 x 150 cm)
Cette ponte particulière est souvent observée par des plongeurs.
Martinique, côte caraïbe
24/02/2007


Une ponte à la Guadeloupe
Une ponte bien colorée donc certainement récente. On remarque quelques interruptions de la coloration : absence d'embryons ?
Sous une large plaque de Sargasses, la piscine (ilêts Pigeons), réserve Cousteau, Malendure, Guadeloupe (971), 3 m sous le mouillage du spot
05/2021


Une ponte sur le platier
Vue rapprochée d'une masse d'œufs. Ces derniers sont bien colorés, la ponte est probablement récente.
Sur le platier de Ngouja au sud de l'île, Mayotte (97620)
08/11/2018


Vieille ponte en pleine eau
La coloration rouge des embryons a disparu et le double cordon d'œufs semble se détacher du cylindre de mucus. Cette ponte est certainement vieille de plusieurs jours. Les embryons ont éclos. Il ne reste donc plus que le manchon gélatineux qui se délite.
Wadi Lahami, Marsa Alam, mer Rouge, Egypte, 3 m
21/11/2010


De plus près
Gros plan sur la ponte précédente. Les deux rangées d'œufs enroulés autour de la masse de mucus sont bien visibles.
Sur le platier de Ngouja au sud de l'île, Mayotte (97620)
08/11/2018


Une ponte particulière
Les embryons sont encore présents dans la partie colorée et dans la partie blanche, les jeunes sont partis.
Cap enragé, Martinique, 1 m
17/07/2007


Vu de très près
Les embryons en cours de développement sont visibles
Pointe des nègres, Martinique, 1 m
05/03/2007


Vieille ponte sur le fond
Cette ponte repose sur le fond. Elle ne devrait pas tarder à se désagréger.
Wadi Lahami, Marsa Alam, mer Rouge, Egypte,15 m
21/11/2010

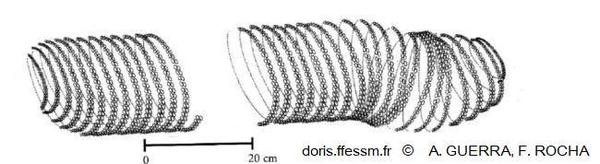
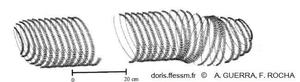
Schéma d'une ponte de Thysanoteuthis rhombus
Ce schéma montre bien les deux rangées parallèles des œufs enroulés autour du cylindre de mucus.
Schéma extrait de Guerra A. et Rocha F. 1997, page 127, figure 2 A.
Reproduction de documents anciens
1997
Rédacteur principal : Yves MÜLLER
Vérificateur : Laure BONNAUD-PONTICELLI
Responsable régional : Yves MÜLLER
Bello G., 2009, On the presence of Thysanoteuthis rhombus (Cephalopoda: Thysanoteuthidae) in the Adriatic Sea, Marine Biodiversity Records, 2.
Biagi V., Bello G., 2009, Occurrence of an egg mass of Thysanoteuthis rhombus (Cephalopoda : Teuthida) in the strait of Messina (Italy) locus typicus of the species, Bollettino. Malacologico, 45, 35-38 (suppl).
Boletzky S. von., 1998, Cephalopod eggs and egg masses, Oceanography and Marine Biology, an Annual Review, 36, 341–371.
Bruggen A.C. van., 1962, Notes on South African marine mollusca 6. Thysanoteuthis rhombus (Cephalopoda) in Algoa Bay, Annals of the Natal Museum, 15(21), 267-271.
Guerra A., Rocha F., 1997, On a floating egg mass of the diamond shaped squid Thysanoteuthis rhombus (Cephalopoda: Thysanoteuthidae) in the western Mediterranean, Iberus, 15(1), 125-130.
Guerra A., Gonzalez A.F., Rocha F.J., Sagarminaga R., Canadas A., 2002, Planktonic egg masses of the diamond-shaped squid Thysanoteuthis rhombus inn the eastern Atlantic and the Mediterranean sea, Journal of Plankton Research, 24(4), 333-338.
Mangold K., Boletzky S. von., 1987, CÉPHALOPODES (SEICHES, CALMARS ET POULPES/PIEUVRES) in Fischer, W., M.-L. Bauchot et M. Schneider (rédacteurs), -1987 FICHES FAO D’IDENTIFICATION DES ESPÈCES POUR LES BESOINS DE LA PÊCHE. (RÉVISION 1). MÉDITERRANÉE ET MER NOIRE. ZONE DE PÊCHE 37. VOLUME I. VÉGÉTAUX ET INVERTÉBRÉS. Publication préparée par la FAO, résultat d’un accord entre la FAO et la Commission des Communautés Européennes (Projet GCP/INT/422/EEC) financée conjointement par ces deux organisations. Rome, FAO, 760 p.
Miyahara K., Fukui K., Nagahama T., Ohatani T., 2006, First record of planktonic egg masses of the diamond squid, Thysanoteuthis rhombus Troschel, in the sea of Japan, Plankton & Benthos Research, 1(1), 59-63.
Miyahara K., Fukui K., Ota T., Minami T., 2006, Laboratory observations on the early life stages of the diamond squid Thysanoteuthis rhombus, Journal of Molluscan Studies, 72(2), 199–205.
Naef A., 1921-23, Cephalopoda. Fauna e Flora de Golfo di Napoli, Monograph, N°35, 863 pp.
Nigmatullin C.M., Arkhipkin A.I., Sabirov R.M., 1991, Structure of the reproductive system of Thysanoteuthis rhombus (Cephalopoda: Oegopsida), Journal of Zoology, 224, 271-283.
Nigmatullin C.M., Arkhipkin A.I., Sabirov R.M., 1995, Age, growth and reproductive biology of diamond-shaped squid Thysanoteuthis rhombus (Oegopsida: Thysanoteuthidae), Marine Ecology Progress Series, 124, 73-87.
Nigmatullin C.M., Arkhipkin A.I., 1998, A review of the biology of the diamondback squid, Thysanoteuthis rhombus (Oegopsida: Thysanoteuthidae), in: Okutani, T.,1998, Contributed papers to International Symposium on Large Pelagic Squids, July 18-19, 1996, for JAMARC's 25th anniversary of its foundation, Japan Marine Fishery Resources Research Center: Tokyo. 269 pp..
Nishimura S., 1966, Notes on the occurrence and biology of the oceanic squid, Thysanoteuthis rhombus Troschel, Japan Publications of the Seto marine biological laboratory, 14(4), 327-349.
Pérez A.E., Elena R.R., Gonzalez Gonzalez A.F., Guerra Sierra A., 2012, On the occurrence of egg masses of the diamond-shaped squid Thysanoteuthis rhombus Troschel, 1857 in the subtropical eastern Atlantic (Canary Islands), A potential commercial species?, Zookeys, 222, 69-76.
Troschel F. H.,1857, Bemerkungen über die Cephalopoden von Messina, Archiv für Naturgeschichte, 23, 41-76.
Wakabayashi T., Tsuchiya K., Segawa S., 2005, Morphological changes with growth in the paralarvae of the diamondback squid Thysanoteuthis rhombus Troschel, 1857, Phuket marine Biological Center of research Bulletin, 66, 167-174.
La page de Thysanoteuthis rhombus dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.














")

