

Coquille ovale, fuselée, globuleuse
20 à 35 mm de hauteur en moyenne
Callosité columellaire bien développée
Couleur de fond beige clair à brun foncé
Nombreuses flammules longitudinales sur la coquille
Manteau blanchâtre moucheté
Nasse renflée, nasse polie, nasse variable, nasse changeante, nasse-ceinture, noisette de Méditerranée
Mutable nassa, changeable nassa (GB), Lumachino di mare (I), Mugarida lisa (E), Glatte Netzreusenschnecke, Wandelbare Reusenschnecke (D)
Buccinum mutabile Linnaeus, 1758
Nassa mutabilis (Linnaeus, 1758)
Nassarius mutabilis (Linnaeus, 1758)
Sphaeronassa mutabilis (Linnaeus, 1758)
Buccinum gibbum Bruguière, 1789
Buccinum foliorum Gmelin, 1791
Buccinum tessulatum Gmelin, 1791
Buccinum jaspideum Link, 1807
Buccinum foliosum W. Wood, 1818
Nassa mediterranea Risso, 1826
Buccinum rufulum Kiener, 1834
Nassa gibba M.E. Gray, 1850
Nassa ebenacea Gennari, 1866
Sphaeronassa globulina Locard, 1886
Nassa helvetica Peyrot, 1925
Sphaeronassa adriatica, Coen, 1933
Sphaeronassa deformis, Coen, 1933
Sphaeronassa umbilicata Coen, 1933
Méditerranée, mer Noire et Afrique de l'Ouest
Zones DORIS : ● Europe (côtes françaises), ○ [Méditerranée française]Cette espèce est largement répandue dans tout le bassin méditerranéen, mer Noire comprise. Sa répartition s’étend sur les côtes occidentales de l'Afrique jusqu’en Mauritanie.
Cette nasse vit en eau peu profonde, entre 2 et 15 m de profondeur, sur les fonds sablo-vaseux, parfois dans les herbiers de zostères, de l’étage infralittoral*. Chasseur nocturne, elle vit enfouie dans le sable durant la journée.
La coquille de ce gastéropode Nassariidé est ovale, fuselée avec un dernier tour globuleux qui occupe pratiquement les 2/3 de la hauteur totale du test*.
Sa taille varie entre 18 et 38 mm de hauteur pour une largeur moyenne de 23 mm.
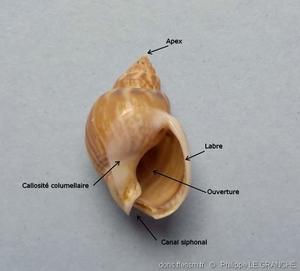
Les tours de spire* sont séparés par une suture* bien distincte. L’ouverture est ovale avec une profonde échancrure siphonale dans sa partie antérieure. Le canal siphonal* est observable postérieurement, il est court et replié dorsalement. Une callosité columellaire* est plus ou moins développée selon l’âge de l’individu sur la partie ventrale du dernier tour. Le labre* est arqué avec des plis sur son bord interne.
De fines lignes axiales et concentriques se croisent formant un treillis sur les 2 premiers tours. On distingue également de légères lignes concentriques à la base du dernier tour de spire.
Sa couleur est assez variable allant du beige clair au brun foncé avec de nombreuses flammules longitudinales généralement plus foncées.
Un petit opercule* corné onguiforme*, situé à l'extrémité de la queue, obstrue l’entrée de la coquille quand l’animal est rétracté. Le manteau* du mollusque est blanchâtre, moucheté de points et de petits traits noirs et blancs. Les tentacules* sont longs et portent des yeux à leur base du côté externe.
Le siphon* est allongé et l’extrémité postérieure du pied* se termine par deux très fines expansions en fourche.
Tritia corniculum : sa taille est plus petite, ne dépassant pas 20 mm. Sa coquille est plus allongée et son dernier tour moins globuleux. Sa couleur est plus foncée et le labre dentelé. Elle n’est présente qu’en Méditerranée et en Bretagne nord.
Tritia nitida : si la hauteur est à peu près comparable, la différence se remarque par un relief particulier de sa coquille avec une ornementation en varices et des lignes spiralées.
La nasse renflée est un mollusque macrophage* et nécrophage* ; c’est donc un excellent éboueur de la mer. Elle se déplace très vite et peut seule ou en groupe dépecer une proie morte ou blessée (poisson, crustacé, autre mollusque bivalve) ou tout cadavre d’animal à proximité de son habitat sablo-vaseux dans lequel elle passe son temps enfouie.
Carnivore charognard, cet animal détecte l'odeur de cadavres d'animaux de très loin à l'aide de son siphon*. Ce dernier possède des récepteurs sensoriels ou chémorécepteurs* particulièrement bien développés et performants sur plusieurs dizaines de mètres lui permettant de repérer rapidement et à distance ses proies potentielles. Il remplit un rôle de premier ordre de nettoyeur dans la chaîne alimentaire.
Cette espèce à sexes séparés est dite gonochorique*. Après copulation, la fécondation est interne. Après la fécondation, qui se situe entre la fin de l’hiver et le printemps (mars – avril), la femelle dépose des capsules ovigères* transparentes en forme d’urnes. Ces capsules sont hérissées de crêtes et d’épines (ce seraient les capsules les plus finement décorées des Nassariidés). Chaque capsule est attachée au support (algue, feuille de phanérogame*, tube de polychète, pierre,…) par sa base. Les capsules sont disposées en rangées.
Souvent les femelles se regroupent pour pondre, d’où la présence d’un grand nombre de capsules à certains endroits. Chaque capsule contient 9 à 27 œufs (16-21 en moyenne) et est fermée à son sommet par un bouchon muqueux. Ce dernier disparaît à l’éclosion des larves*. Des observations ont été faites sur les côtes catalanes espagnoles où Tritia mutabilis utilisait volontairement les œufs de la seiche commune, Sepia officinalis, comme support.
Les larves* planctotrophiques* ont une longue vie pélagique*, près de deux mois pendant lesquels elles vont se métamorphoser en plusieurs stades, trochophore* puis véligère*, avant de tomber sur le fond et se transformer en petite nasse juvénile identique à l’adulte.
Les coquilles vides de Tritia mutabilis sont souvent occupées par des pagures (comme Paguristes eremita).
Cette nasse a pour principal prédateur les étoiles de mer notamment les étoiles-peignes (Astropecten spp.).
La nasse possède un intéressant mécanisme de fuite : lorsqu’une étoile de mer s’approche assez près d’elle pour la toucher, elle bascule sa coquille vers l’avant et dégage son pied du substrat*, de telle façon que l’ouverture de la coquille se trouve alors vers le haut. Elle se retourne ensuite en contractant son pied afin de reprendre contact avec le fond. Elle se catapulte ainsi une dizaine de fois en zigzag pour fuir son prédateur. Cette méthode lui permet de s’échapper plus rapidement qu’en rampant.
Cette espèce, très commune, est une ressource importante pour la pêche artisanale principalement en Italie (Sicile, mer Adriatique, golfe de Gênes) mais également depuis 2011 dans le golfe du Lion, en particulier au large du Grau-du-Roi, aux Saintes-Maries-de-la-Mer, à Agde et à Port-la- Nouvelle. Elle est récoltée par de petites unités à l’aide de dragues ou de chaluts de fonds. Elle peut être consommée crue ou en conserve.
On a constaté depuis des années une constante diminution des prises à cause de la surpêche, de la prédation des capsules ovigères par les pagures et par les effets négatifs sur la reproduction dus aux peintures antifouling.
Pour y pallier, des essais de récifs artificiels de forme pyramidale ont été tentés avec succès en mer Adriatique, dans la région des Abruzzes (Italie). Déposés sur des fonds sableux vers 6 m de profondeur, ils offrent aux capsules un substrat idéal pour se fixer.
Selon certaines études, cette espèce s’enfouirait dans le sédiment quand la température de l’eau augmente.
Nasse : le dessin treillissé de la coquille fait penser au piège en osier composé de lames tressées de bambou, roseaux ou bois, ou en métal recouvert d'un filet de pêche.
renflée : comme le dernier tour globuleux de la coquille.
Tritia : Risso, en 1826, n’a pas donné d’explication. Peut-être Triteia une nymphe marine fille du dieu marin Triton dans la mythologie grecque.
mutabilis : mot latin = sujet au changement, variable, en relation avec la variabilité de la coloration de la coquille.
Numéro d'entrée WoRMS : 876640
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Gastropoda | Gastéropodes | Mollusques à tête bien distincte, le plus souvent pourvus d’une coquille dorsale d’une seule pièce, torsadée. La tête porte une ou deux paires de tentacules dorsaux et deux yeux situés à la base, ou à l’extrémité des tentacules. |
| Sous-classe | Caenogastropoda | Caenogastropodes | |
| Ordre | Neogastropoda | Néogastéropodes | Coquille avec canal siphonal bien développé. Un repli du manteau forme un tube extensible : le siphon. La plupart sont des prédateurs ou nécrophages. Tous marins sauf le genre Clea. |
| Famille | Nassariidae | Nassariidés | Coquille solide ovoïde à globuleuse, de petite taille (4-75mm); sculpture spiralée, axiale ou dans les deux sens (treillissage). Ouverture circulaire-ovale, souvent avec une découpe circulaire et un sillon (siphon anal) en haut, généralement tronquée en bas et avec un canl siphonal court ou un sillon recourbé en arrière. Labre à bord interne denté. Columelle en général plissée, callosité columellairesouvent large (spécimens adultes). Opercule corné, arrondi-triangulaire ou en forme de griffe, à nucléus terminal et souvent à bord dentelé. Lindner 2011:111. |
| Sous-famille | Nassariinae | Nassariinés | |
| Genre | Tritia | ||
| Espèce | mutabilis |
") Gastéropodes à coquille unique : escargots subaquatiques et assimilés (ex. Prosobranches et Hétérobranches/Pulmonés)
Gastéropodes à coquille unique : escargots subaquatiques et assimilés (ex. Prosobranches et Hétérobranches/Pulmonés)

Flammules plus foncées sur fond clair
La couleur de la coquille est assez variable allant du beige clair au brun foncé avec de nombreuses flammules longitudinales généralement plus foncées.
Limnos, Grèce, mer Egée, 2 m
29/06/2012
Gastéropodes à coquille unique : escargots subaquatiques et assimilés (ex. Prosobranches et Hétérobranches/Pulmonés)

Dernier tour globuleux
La coquille de ce gastéropode Nassariidé est fuselée avec un dernier tour globuleux qui occupe pratiquement les 2/3 de la hauteur totale du test.
Limnos, Grèce, mer Egée, 2 m
29/06/2012


Tête en gros plan
Les deux tentacules portent à leur base un œil noir et le siphon est bien visible.
Port-Camargue (30), 2 m , de nuit
28/03/2019


Siphon
Cet animal détecte l'odeur de cadavres d'animaux de très loin à l'aide de son siphon.
Cros-de-Cagnes (06), 10 m, de nuit.
07/03/2020


Manteau blanchâtre moucheté
Le manteau du mollusque est blanchâtre, moucheté de points et de petits traits noirs et blancs.
Cros-de-Cagnes (06), 10 m, de nuit.
07/03/2020


Suture bien distincte
Les tours de spire sont séparés par une suture bien distincte.
Cros de Cagnes (06), 6 m
01/11/2017


Juvénile
Ce juvénile arbore des teintes de coquille et de manteau plus claires.
Karpathos, archipel du Dodécanèse, Grèce, 3 m
24/09/2011


En chasse de nuit
Cette nasse est un mollusque carnivore charognard nocturne.
Les Remparts, Antibes (06), 12 m, de nuit.
23/08/2013


Dessus-dessous
Selon leur activité, on peut observer simultanément des individus sur le sable, ou quasiment entièrement enfouis sous sa surface, laissant dépasser l'extrémité de leur siphon.
Cros-de-Cagnes (06), 5 m, de nuit
13/05/2017


Pied fourchu
Cette photo nous permet de bien remarquer l'extrémité fourchue du pied de ce gastéropode.
Cros-de-Cagnes, (06), 5 m, de nuit
05/08/2011


Opercule
Un petit opercule corné onguiforme situé à l'extrémité de la queue obstrue l’entrée de la coquille quand l’animal est rétracté.
Limnos, Grèce, mer Egée, 2 m
29/06/2012


Macrophage et nécrophage
La nasse renflée se déplace très vite et peut seule ou en groupe dépecer une proie morte ou blessée (poisson, crustacé, autre mollusque).
Cros-de-Cagnes (06), 6 m, de nuit
19/10/2014


Prédateur
Cette nasse a pour principal prédateur les étoiles de mer notamment les étoiles-peignes (Astropecten spp.).
Limnos, Grèce, mer Egée, 2 m
29/06/2012


Nettoyage ou prédation ?
Rouget en fin de vie prêt à être dévoré ou nettoyage des écailles ?
Cros-de-Cagnes (06), 6 m, de nuit.
07/03/2020


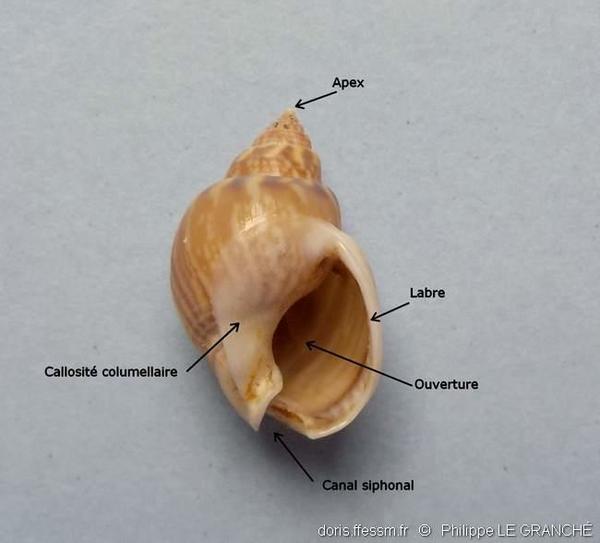
Pour mieux comprendre
Différentes parties d’une coquille de 20 x 13 mm.
Méditerranée
29/01/2018


Couleur variable
La couleur de cette nasse est variable, allant du beige clair au brun foncé avec de nombreuses flammules longitudinales généralement plus foncées.
Extrait de : Iconographie der schalentragenden europäischen Meeresconchylien, vol. II, Cassel, gravure XXII de W. Kobelt
Reproduction de documents anciens
1887

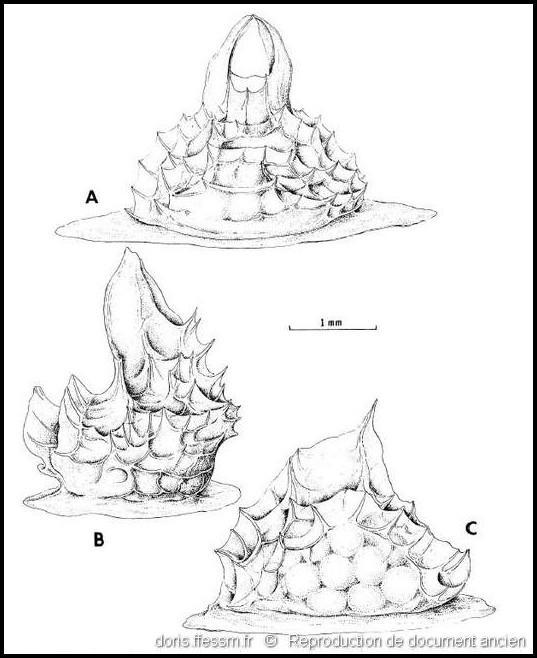
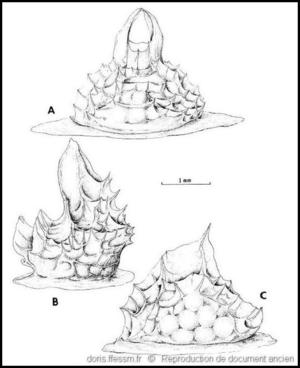
Capsules ovigères
A : côté opposé à l’ouverture de sortie des œufs montrant les projections apicales qui s’arquent sur le bouchon muqueux, B : vue latérale, C : côté avec l’ouverture de sortie des œufs d’une capsule dépourvue de saillies apicales.
Dessin de C.N. D’Asaro (fig 19) extrait de l'ouvrage " Gunnar Thorson’s world-wide collection of prosobranch egg capsules: Nassariidae". D’après un échantillon recueilli à proximité de Naples (Italie).
Reproduction de documents anciens
1993


Capsules in-situ
Ces capsules ont été déposées dans une coquille d'huitre morte dans une zone vaseuse.
Fos-sur-Mer (13), 5 m
24/02/2024
Rédacteur principal : Philippe LE GRANCHÉ
Vérificateur : Yves MÜLLER
Responsable régional : Philippe LE GRANCHÉ
Aissaoui C., Galindo L.A., Puillandre N., Bouchet Ph., 2017, The nassariids from the Gulf of Gabès revisited (Neogastropoda, Nassariidae), Marine Biology Research, 13, 4, 370-389.
Caprioli R., Giansante C., 2018, Preliminary investigation on the use of artificial substrates to favor Tritia mutabilis (Linnaeus, 1758) spawning in Central Adriatic Sea: a possible contribution to stock maintenance, Acta Adriatica (I), 59(1), 141-148.
D’Asaro C.N., 1993, Gunnar Thorson’s world-wide collection of prosobranch egg capsules: Nassariidae, Ophelia, 38(3), 149-215.
Kohn A.J., 1961, Chemoreception in gastropod molluscs, American zoologist, 1, 291-308.
La page de Tritia mutabilis sur le site de référence de DORIS pour les mollusques : MolluscaBase
La page de Tritia mutabilis dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.



















")

