

Algue brune pouvant atteindre 50 cm
Touffes laineuses composées de filaments très fins
Filaments très ramifiés
Filaments enroulés à la manière d’une corde près de la base
Au microscope : filaments unisériés
Au microscope : présence de cystes uniloculaires ou pluriloculaires intercalaires
Conferva littoralis Linné, 1753
Ceramium littorale (Linnaeus) C.Agardh 1817
Ectocarpus littoralis (Linnaeus) Lyngbye 1819
Ectocarpus brachiatus (Smith) S.F.Gray 1821
Ectocarpus brachiatus (C.Agardh) C.Agardh 1824
Ectocarpus ferrugineus C.Agardh 1824
Ectocarpus rufus (Roth) C.Agardh 1824
Ectocarpus compactus (Roth) C.Agardh 1828
Ectocarpus cruciatus C.Agardh 1828
Lyngbya littoralis (Linnaeus) Dillwyn ex Gaillon 1828
Ectocarpus castaneus Kützing 1843
Ectocarpus ramellosus Kützing 1845
Ectocarpus rutilans Kützing 1845
Ectocarpus subverticillatus Kützing 1845
Spongomorpha castanea (Kützing) Kützing 1845
Spongomorpha ferruginea Kützing 1845
Ectocarpus firmus (C. Agarth) J.Agarth 1848
Compte tenu de la grande variété phénotypique* de l’espèce, les synonymes* du nom scientifique actuel sont trop nombreux pour être tous cités ici.
Atlantique Nord-Est
Zones DORIS : ● Europe (côtes françaises), ○ [Atlantique Nord-Est, Manche et mer du Nord françaises], ○ [Méditerranée française], ● Atlantique Nord-OuestCette espèce est cosmopolite en zones tempérées. Elle est présente dans l'Atlantique Nord-Est, en Méditerranée (introduite en Méditerranée française) et en Adriatique. L’espèce est signalée sur les côtes du Canada et des États-Unis
Pylaiella littoralis est majoritairement rencontré en épiphyte* de Fucus serratus, donc au niveau du médiolittoral* moyen. Cette espèce peut parfois être épiphyte d’autres algues, notamment du genre Fucus, ou se fixer directement sur la roche. L’espèce est euryhaline* et se développe très bien en estuaire. Elle est sensible à l’agitation des eaux et préfère donc les zones faiblement à moyennement exposées. Elle tolère très bien les eaux enrichies, qui peuvent conduire sous certaines conditions à des proliférations estivales.
Cette algue brune forme des touffes laineuses brunâtres à verdâtres. Le thalle* est formé de filaments unisériés* très fins abondamment ramifiés, dont la longueur atteint fréquemment 20 cm. Certains thalles peuvent être significativement plus longs (40 cm et plus). Les ramifications peuvent être unilatérales ou opposées.
Les filaments présentent la particularité de s’enrouler les uns autour des autres à la base du thalle, à la manière d’une corde, ce phénomène étant facilité par la présence de certaines branches très fines.
Au microscope, on peut constater que les filaments sont unisériés*. Les cellules, d’un diamètre maximal de 50 µm, peuvent être plus ou moins allongées. Elles possèdent de nombreux plastes* discoïdes de couleur brune et unseul pyrénoïde* sans disposition particulière perceptible au sein des cellules..
Les thalles fertiles produisent des cystes* intercalaires (entre des cellules végétatives) uniloculaires* ou pluriloculaires.
Les cystes uniloculaires peuvent parfois être aussi terminaux.
De manière occasionnelle, il est possible d’observer des paires de cystes uniloculaires, suite à une division longitudinale des cellules concernées (ces divisions longitudinales peuvent également être observées sur les cellules végétatives).
Pylaiella littoralis, qui est aujourd’hui reconnu comme un complexe d’espèces, présente une grande plasticité phénotypique, ce qui a conduit les auteurs à proposer de nombreux noms d’espèces ou de sous-espèces (plus de 90 !). Des tentatives d’analyse informatique des différents morphes* (voir Bolton 1979) avaient conduit à la conclusion que les différentes variations morphologiques constatées sur le terrain ne pouvaient être rapportées simplement à des espèces différentes. Ce n’est que l’émergence des techniques moléculaires modernes qui a permis de clarifier les choses (si tant est que les conclusions tirées de ces techniques puissent être considérées comme définitives…).
De très nombreuses algues Ectocarpales se présentent sous la forme de filaments fins brunâtres semblables. Le « tressage » de la partie proximale des filaments est un critère distinctif pertinent. La détermination certaine nécessitera un examen au microscope d’individus fertiles.
Les thalles de Pylaiella littoralis atteignent par ailleurs des longueurs rarement rencontrées chez les autres Ectocarpales.
Comme toutes les algues, Pylaiella littoralis est un organisme autotrophe* photosynthétique*. L'algue tire son énergie de la lumière solaire, et grâce à l'absorption d'eau, de dioxyde de carbone et des sels minéraux dissous dans l'eau, elle fabrique les matières organiques nécessaires à son développement.
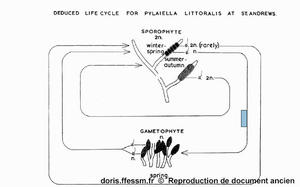
Pylaiella littoralis se reproduit selon un cycle digénétique* haplodiplonte* isomorphe* mais avec de nombreuses variantes.
Les cystes* uniloculaires* ou pluriloculaires libèrent des cellules mobiles, dotées de deux fouets latéraux de longueur inégale. Les cystes uniloculaires libèrent des spores* haploïdes* (128 ou 256 par cellule) qui vont se fixer et donner naissance à des thalles* haploïdes (gamétophytes*).
La première division cellulaire dans les cystes uniloculaires est méïotique, et conduit donc à une réduction du nombre de chromosomes dans chaque cellule-fille.
Les cystes pluriloculaires libèrent des cellules plus complexes, car selon les cas elles peuvent être des gamètes* sexués (haploïdes) ou des spores (diploïdes*). Les gamètes peuvent être fécondés par un autre gamète et donner naissance à un thalle diploïde (sporophyte*), ou se fixer directement et donner naissance à un nouveau gamétophyte. Les spores diploïdes donneront directement naissance à un sporophyte.
Les thalles contenant des cystes uniloculaires sont donc des sporophytes, tandis que les thalles contenant des cystes pluriloculaires sont des gamétophytes. Certains sporophytes comportent des cystes pluriloculaires. Ces cystes libéreront des spores qui se développeront directement, sans fécondation.
Le cycle complet de Pylaiella littoralis passe par l’alternance de ces deux types de thalles, à l’allure identique. Dans une population donnée, les sporophytes sont généralement plus nombreux que les gamétophytes.
Il semblerait que certaines souches de cette espèce ne se reproduisent que de façon végétative. Il est à noter, en particulier, que des travaux modernes (Müller & Stächer, 1989) ont remis en cause l’existence de gamétophytes (les thalles ne comportant que des cystes pluriloculaires, considérés comme des gamétophytes, pouvant présenter des cystes uniloculaires lorsque les conditions de culture sont modifiées).
Dans les eaux de la Manche, le pic de fertilité est observé à la mi-octobre, peu avant le déclin des populations durant la saison froide.
Dans certaines zones, on peut observer une évolution du type de cystes en fonction de la saison (voir figure issue des travaux de George Russel). Des thalles portant des cystes pluriloculaires sont plutôt rencontrés en été et en automne, alors que des thalles portes des cystes uniloculaires sont rencontrés au printemps. Ces observations ne sont cependant pas généralisables à d’autres zones géographiques.
Pylaiella littoralis vit en épiphyte* sur de nombreuses algues, mais l’association la plus fréquente est celle avec Fucus serratus. On peut également la trouver sur d’autres espèces de Fucus, notamment en zones abritées, sur Ascophyllum nodosum ou sur Chorda filum.
On peut observer une succession des plantes hôtes au cours de l’année. Margery Knight (1923) avait ainsi observé que Ascophyllum nodosum était principalement hôte au printemps, puis Fucus vesiculosus et enfin Fucus serratus.
De manière intéressante, il a été constaté que les cellules reproductrices sont différentes selon l’algue support. Près de 90 % des thalles* fertiles ayant pour support Ascophyllum nodosum présentent des cystes* pluriloculaires*. Ce type de cystes n’est au contraire présent que sur 2 % des thalles sur Fucus spiralis et 15 % des thalles sur Fucus vesiculosus (ces données concernent les observations faites par Margery Knight sur des populations de l’île de Man).
L’espèce peut également se développer directement sur la roche. Une forme totalement libre a également été observée sur les côtes du Massachusetts, les thalles formant alors des boules dérivant au gré des courants.
Pylaiella littoralis peut être parasité par les oomycètes Anisolpidium rosenvingei et Eurychasma dicksonii et par le champignon Chytridium polysiphoniae. Ces parasites* ne sont pas spécifiques à cette algue mais peuvent coloniser de nombreuses autres algues brunes. Ils n'induisent pas d'effets biologiques significatifs sur les hôtes. Anisolpidium rosenvingei parasite uniquement les cellules reproductrices de l’algue et le taux de parasitisme est maximal au pic de reproduction de l’espèce. Les deux autres espèces parasitent les cellules végétatives. Eurychasma dicksonii serait responsable de l’apparition de thalles libres (voir supra) en fragilisant le thalle au niveau de ses sporanges* lorsque ceux-ci ont été vidés.
Des études récentes ont montré que, sur les côtes du Maine où Pylaeilla littoralis est essentiellement présente sous forme non fixée, l’amphipode Gammarus tigrinus consomme principalement cette algue. Les fèces de cette espèce comportent de nombreux fragments de thalle encore aptes à survivre. Le crustacé contribue ainsi à la propagation de l’algue.
L’espèce peut devenir très envahissante l’été. La température, l’éclairement et la pollution azotée favorisent ces proliférations.
Si l’espèce est clairement euryhaline*, il a été démontré qu’il existe différents écotypes*, plus ou moins adaptés aux faibles salinités. Ces facultés d’adaptation sont d’origine génétique.
La présence de cette espèce en Méditerranée occidentale (étang de Thau) est considérée comme étant d’origine anthropique* (sans doute introduite de l’Atlantique ou du Pacifique Nord avec des huîtres).
Les extraits de Pylaiella littoralis ont un effet marqué sur certaines cellules cancéreuses. Cette algue pourrait ouvrir une voie prometteuse dans le traitement des cancers colorectaux, comme d’autres algues brunes.
Algue filamenteuse de La Pylaie : le nom français proposé reprend l’hommage à M. de La Pylaie.
Pylaiella : ce nom de genre Pylaiella a été créé en 1823 par le naturaliste, et officier français, Jean-Baptiste Geneviève Marcellin Bory de Saint-Vincent (1778-1846) en hommage au botaniste et explorateur français Auguste Jean-Marie Bachelot de La Pylaie (1786-1856).
littoralis : le nom d’espèce littoralis provient du latin [litoralis] = du rivage. Cette espèce est rencontrée sur l'estran*.
Numéro d'entrée WoRMS : 157188
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Ochrophyta | Ochrophytes | ou Hétérokontes, ou Straménopiles: présence d'un stade unicellulaire à 2 flagelles, un lisse et un à poils tubulaires. |
| Classe | Phaeophyceae | Phéophycées | Algues brunes. |
| Sous-classe | Fucophycidae | ||
| Ordre | Ectocarpales | Ectocarpales | |
| Famille | Acinetosporaceae | Acinetospracées | |
| Genre | Pylaiella | ||
| Espèce | littoralis |
") Algues brunes (Phéophycées)
Algues brunes (Phéophycées)

Vue générale
Le thalle* est ici en épiphyte* de Fucus spiralis. On voit nettement l’aspect laineux qui s’en dégage lorsqu’il est immergé.
Estran, Agon Coutainville (50), Manche Ouest
16/09/2012
Algues brunes (Phéophycées)

Vue de plusieurs thalles épiphytes de Fucus serratus
Cette photographie permet de voir la physionomie typique des thalles* de Pylaiella littoralis, formés de filaments bruns qui forment une sorte de tresse à leur base.
Photographie prise dans une mare médiolittorale*, Binic (22)
21/08/2025

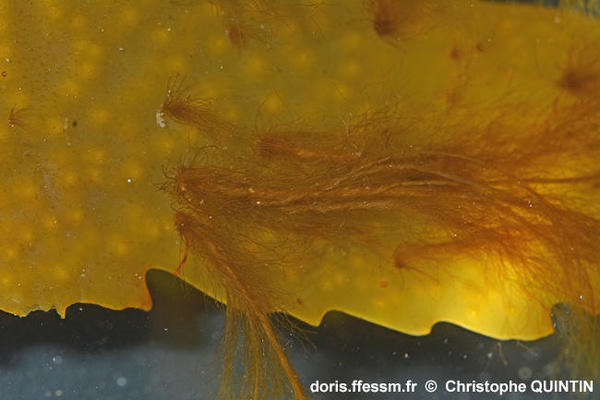
Vue rapprochée de plusieurs thalles épiphytes de Fucus serratus
On observe ici la physionomie typique des thalles* de Pylaiella littoralis, formés de filaments bruns qui forment une sorte de tresse à leur base.
Photographie prise dans une mare médiolittorale, Binic (22)
21/08/2025

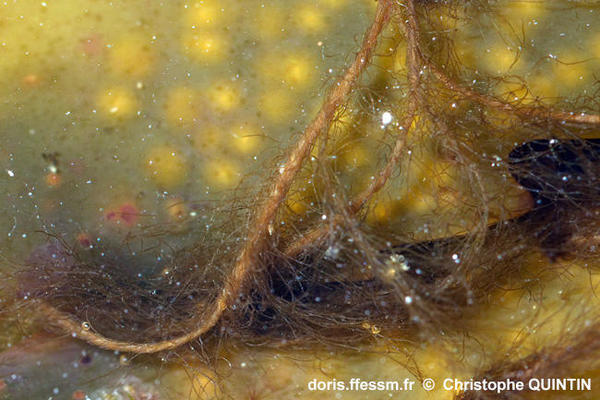
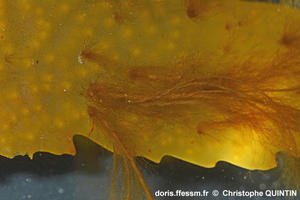
Vue rapprochée de la partie proximale (basale) d’un thalle
On voit nettement les tresses qui se forment à la base du thalle*.
Photographie prise dans une mare médiolittorale*, Binic (22)
21/08/2025


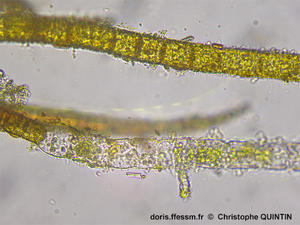
Vue au microscope de la partie proximale d’un thalle
Cette microphotographie permet de voir la manière dont les filaments s’enroulent à la base du thalle*, à la manière d’une corde. La composition unisériée* des filaments est ici clairement visible.
Photographie prise au microscope, Binic (22)
21/08/2025


Vue au microscope de la partie courante d’un thalle
Les filaments de Pylaiella littoralis sont unisériés et ramifiés.
Photographie prise au microscope, Binic (22)
21/08/2025


Vue au microscope d’une ramification
On peut observer les filaments unisériés et une ramification.
Photographie prise au microscope, Binic (22)
21/08/2025


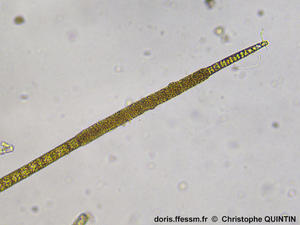
Vue au microscope d’un filament
Cette microphotographie permet de voir les filaments unisériés, chaque cellule étant dotée de plastes bruns discoïdes. On peut voir que les cellules sont de longueur variable.
Photographie prise au microscope, Binic (22)
21/08/2025

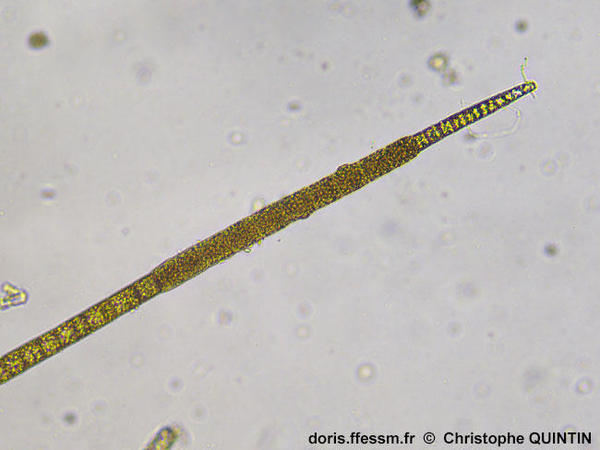
Vue des structures reproductrices asexuées
Cette microphotographie permet de voir les cystes* uniloculaires* intercalaires, qui contiennent des spores*. Ces cellules sont plus larges et plus sombres que les cellules végétatives. Elles s’arrondissent en mûrissant.
Photographie prise au microscope, Binic (22)
12/10/2025


Vue des structures reproductrices asexuées
On constate la présence des cystes* uniloculaires* intercalaires, qui contiennent des spores*. Ces cellules sont plus larges et plus sombres que les cellules végétatives. Elles s’arrondissent en mûrissant.
Photographie prise au microscope, Binic (22)
12/10/2025


Vue des structures reproductrices asexuées après émission des spores
On voit ici, sur la branche du bas, d’anciennes cellules comportant des spores*, reconnaissables au fait qu’elles sont plus larges que les cellules adjacentes, qui se sont vidées. Ces cellules vides constituent des zones de fragilité du thalle, qui se découpera probablement à ce niveau lors de la prochaine tempête. L’excroissance latérale observée sur l’une des cellules vides pourrait correspondre au tube de sortie de l’oomycète parasite Anisolpidium rosenvingei.
Photographie prise au microscope, Binic (22)
12/10/2025


Vue de diatomées épiphytes du genre Synedra
Malgré la petitesse de ses rameaux, Pylaiella littoralis peut servir de support à de nombreuses espèces épiphytes*, et notamment des diatomées* comme Synedra sp. ou Grammatophora sp..
Photographie prise au microscope, Binic (22)
12/10/2025

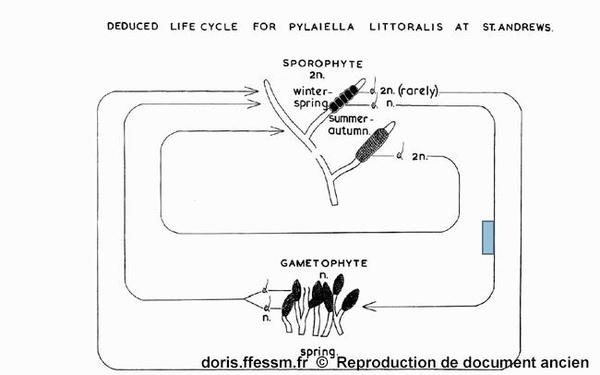
Vue synthétique du cycle de vie de Pylaiella littoralis
Russel G., 1959, The autoecology of Pylaiella littoralis (L.) Kjellm. at St. Andrews, Scotland with additional information on the life cycle of Pylaiella rupincola (Aresh.) Kylin
Reproduction de documents anciens
1959
Rédacteur principal : Christophe QUINTIN
Vérificateur : Marc VERLAQUE
Responsable régional : Yves MÜLLER
Ben Maiz N., 1986, Sur la présence de Pylaiella littoralis (Linnaeus) Kjellman (Phaeophyceae,Ectocarpale) dans l’Etang de Thau (Hérault, France), Rapports et Procès Verbaux des Réunions, Commission Internationale pour l’Exploration Scientifique de la mer Méditerranée, 30 (2), 6.
Bolton J.J., 1979a, Estuarine Adaptation in Populations of Pilayella littoralis (L.) Kjellm. (Phaeophyta, Ectocarpales), Estuarine and Coastal Marine Science, 9, 273-280.
Bolton J.J., 1979b, The taxonomy of Pilayella littoralis (L.) Kjellm. (Phaeophyta, Ectocarpales) in the British Isles, a numerical approach, British Phycological Journal, 14(4), 317-325,
Geoffroy A., Mauger S., de Jode A., Le Gall L., Destombe C., 2015, Molecular evidence for the coexistence of two sibling species in Pylaiella littoralis (Ectocarpales, Phaeophyceae) along the Brittany coast, Journal of Phycology, 51 (3), 480-489, 10.1111/jpy.12291.
Gleason F., Küpper F., Amon J., Picard K., Gachon C., Marano A., Sime-Ngando T., Lilje O., 2011, Zoosporic true fungi in marine ecosystems: a review, Marine and Freshwater Research, 62, 383–393.
Kjellman F.R., 1872, Bidrag till kännedomen om Skandinaviens Ectocarpeer och Tilopterider, Stockholm: Akademisk afhandling Upsala, 1-112, 2 plates.
Knight, M., 1923, Studies in the Ectocarpaceae I The life-history and cytology of Pylaiella littoralis Kjellm., Transactions of the Royal Society of Edinburgh 53, 343-360, Plates 1-6.
Küpper F., Müller D., 1999, Massive occurrence of the heterokont and fungal parasites Anisolpidium, Eurychasma and Chytridium in Pylaiella littoralis (Ectocarpales, Phaeophyceae), Nova Hedwigia, 69, 381-389. 10.1127/nova.hedwigia/69/1999/381.
Miller S., Wilce R., 2024, Grazing of free-living Pylaiella littoralis by the amphipod Gammarus tigrinus, Botanica Marina, 67(1),11-14.
Müller D.G., Stäche B., 1989, Life History Studies on Pilayella littoralis (L.) Kjellman (Phaeophyceae, Ectocarpales) of Different Geographical Origin, Botanica Marina, 32, 71-78.
Russel G., 1959, The autoecology of Pylaiella littoralis (L.) Kjellm. at St. Andrews, Scotland with additional information on the life cycle of Pylaiella rupincola (Aresh.) Kylin, Thèse de doctorat de l’université de Saint Andrews.
Russel G., 1963, A study in populations of Pylaiella littoralis, Journal of the Marine Biological Association of the United Kingdom, 43, 469-483.
Verlaque, M., Ruitton, S., Mineur, F., Boudouresque, C.-F., 2015, CIESM atlas of exotic species of the Mediterranean Macrophytes, Monaco, CIESM Publishers.
West J. A., 1967, Pilayella littoralis F. rupincola from Washington: the life history in culture, Journal of Molecular Evolution, 3, 150-153.
Wilce R., Schneider C., Quinlan A, Bosch K., 1982, The life history and morphology of free-living Pilayella littoralis (L.) Kjellm. (Ectocarpaceae, Ectocarpales) in Nahant Bay, Massachusetts, Phycologia, 21, 336-354.
Ye B., Kim J., Kim M., Jang J., Oh C., Kang D., Qian Z., Jung W., Choi I., Heo S., 2013, Induction of Apoptosis by the Tropical Seaweed Pylaiella littoralis in HT-29 Cells via the Mitochondrial and MAPK Pathways, Ocean Science Journal, 48(4), 339-348.
La page sur Pylaiella littoralis sur le site de référence de DORIS pour les algues : AlgaeBase
La page de Pylaiella littoralis sur le site de l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.










")

