

Algue unicellulaire visible au microscope optique
Diatomée rectangulaire en vue connective
Un tube prolongeant la cellule de chaque côté
Présence d’un velum à chaque extrémité de la cellule
Triceratium brightwellii T. West, 1860
Ditylum inaequale J.W. Bailey ex L.W. Bailey, 1862
Ditylum trigonum L.W. Bailey ex L.W. Bailey, 1862
Cosmopolite sauf dans les eaux polaires
Zones DORIS : ○ [Atlantique Nord-Est, Manche et mer du Nord françaises]Ditylum brightwellii est une diatomée* cosmopolite absente des eaux polaires. Elle se rencontre depuis la surface jusqu’à des profondeurs importantes (470 m) et dans des plages de température très larges (0-29 °C). Il s’agit d’une espèce marine qui supporte moins bien la dessalure (limite basse vers 25 g/L).
Ditylum brightwellii est une espèce planctonique*.
Ditylum brightwellii est une diatomée* centrique. Elle possède une enveloppe externe siliceuse, le frustule*, qui fait l’objet de la description qui suit.
Ditylum brightwellii se présente, en vue connective*, sous la forme d’un rectangle aux angles arrondis, s’achevant aux deux extrémités par un tube, la rimoportula*, dont la longueur peut être légèrement supérieure à la largeur du rectangle. Les deux extrémités du frustule sont par ailleurs bordées par une rangée de courtes épines, qui forment un velum*.
Comme toutes les diatomées, les frustules de Ditylum brightwellii sont constitués de deux valves inégales, l’épithèque (la valve la plus grande) et l’hypothèque, reliées entre elles par une bande siliceuse, le cingulum. Chez Ditylum brightwellii, le cingulum est très long, ce qui donne l’impression que l’épithèque et l’hypothèque ont la même largeur.
En vue valvaire*, Ditylum brightwellii présente une section triangulaire (il existe des formes aberrantes quadrangulaires ou biangulaires), avec quelques crêtes. Ces crêtes correspondent à des lignes plus sombres en vue connective. La zone où débouche le tube central est bordée de nombreuses aréoles* disposées radialement, dont la taille décroit avec leur distance au centre.
Le noyau est central. Les chloroplastes* sont petits et nombreux. La couleur générale de l’algue est brun doré.
La longueur maximale est de 300 µm (130 µm sans les tubes) et la largeur maximale de 100 µm. Le rapport longueur/largeur varie considérablement selon les individus.
Ditylum sol (Grunow) De toni (1894) est une espèce ayant une affinité marquée pour les eaux chaudes mais elle peut être rencontrée dans des eaux plus tempérées. Elle se différencie de Ditylum brigtwellii par la disposition des aréoles* autour du tube central et par le fait qu’elle présente un plus grand nombre de côtes longitudinales. Elle peut par ailleurs être plus large (200 µm) et donc présente un aspect plus carré en vue connective*.
Ditylum brightwellii est une algue autotrophe*, se nourrissant grâce à la photosynthèse*.
Les diatomées* sont des diplontes* (cellules diploïdes*), leur cycle de vie est diplophasique, c'est-à-dire qu'il est constitué essentiellement d'une phase diploïde*, la diplophase.
La phase haploïde* (l'haplophase) est réduite à l'existence des gamètes*.
La reproduction des diatomées peut s’effectuer soit de manière asexuée (multiplication cellulaire végétative), lorsque les conditions de vie sont favorables, soit de manière sexuée.
La multiplication végétative des diatomées est principalement assurée par division cellulaire mitotique, selon un mode très original : après une mitose*, les deux valves du frustule* de l’algue se séparent. Chaque cellule fille conserve une valve de la cellule mère et en sécrète une seconde, qui sera nécessairement une hypothèque, donc plus petite que la valve conservée. Chaque cycle mitotique donne donc naissance à deux individus, l’un de taille similaire à la cellule mère, l’autre plus petit.
En conditions favorables, ce type de reproduction peut être très rapide, une division complète prenant moins d’une journée. La taille moyenne d'une population diminue donc avec le temps.
Il existe une taille minimale seuil au-delà de laquelle peut s’engager une phase de reproduction sexuée qui permettra de restaurer la taille maximale spécifique. La taille minimale seuil varie selon les espèces et est en moyenne de 30 % de la taille de la première cellule ayant engagé un cycle de division mitotique suite à une phase de reproduction sexuée (cellule initiale, voir ci-dessous).
Chez Ditylum brightwellii, il a également été observé une phase de multiplication végétative conduisant à un ré-accroissement de la taille des cellules filles, celles-ci se présentant alors sous une forme différente (section cylindrique et non plus triangulaire).
Pour la reproduction sexuée, les gamètes* mâles et femelles (produits par deux individus différents appelés gamétocystes*) sont générés par une première phase de mitose* (jusqu’à obtenir 16 cellules pour les mâles) puis par méïose*, sur un laps de temps très court (il faut moins de 24 h à un individu pour produire des gamètes). Les petits individus ont tendance à produire préférentiellement des cellules mâles, alors que les individus plus gros donneront soit uniquement des gamètes femelles, soit pour les uns, des mâles, pour les autres, des femelles en proportions identiques. Les individus sexués fournissent deux gamètes pour les cellules femelles (gamétocystes femelles ou oocystes) et 32 à 64 gamètes pour les cellules mâles (gamétocystes mâles ou spermatocystes*). A maturité, le spermatocyste se tord et libère les gamètes mâles gonflant parfois pour faciliter l’ouverture du frustule et leur sortie.
Chez les diatomées centriques, comme Ditylum brightwellii, les gamètes mâles sont munis d’un flagelle* (antérieur chez cette espèce) qui leur permet de rejoindre la diatomée femelle, dans laquelle ils pénètrent grâce à une ouverture produite dans le frustule entre les deux valves. La rencontre entre les gamètes de sexe complémentaire est probablement facilitée par la sécrétion de phéromones* (des ectocarpènes ont été identifiées chez Lithodesmium undulatum, une espèce proche).
La fusion des gamètes mâles et femelles donne naissance à une cellule œuf, qui va s'allonger et se transformer en auxospore*, de flottabilité neutre une fois débarrassée du reste du frustule femelle (mais cette séparation n’est pas obligatoire, auquel cas l’auxospore débutera son développement en restant attaché au frustule maternel). L’auxospore s’entoure d’un abondant mucilage* puis grossit jusqu’à atteindre la taille maximale de l'espèce, avant de se transformer en cellule initiale qui sécrétera alors son frustule siliceux. La cellule initiale permet de régénérer des individus de taille maximale après la diminution de la taille moyenne d'une population, suite aux mitoses successives.
En début de cycle d'auxosporulation, le cytoplasme* de l’auxospore est concentré au milieu de la cellule. Progressivement, les chloroplastes* vont prendre possession de l’ensemble du volume du frustule. Les auxospores n’ont jamais été observées dans les populations naturelles de Ditylum brightwellii, mais sont observées en culture.
Chez Ditylum brightwellii, les individus qui débutent la phase de reproduction peuvent être identifiés par un changement de forme (cellule plus allongée et courbée) et un assombrissement.
Les auxospores peuvent également être utilisées par certaines diatomées pour engager une phase dormante ou de conservation, lorsque les conditions ambiantes deviennent défavorables, mais cela ne semble pas avoir été observé chez Ditylum brightwellii.
Anya Waitel et Paul J. Arrison (1992) ont étudié la reproduction sexuée de Ditylum brightwellii, en milieu naturel et au laboratoire. Ils ont montré que les individus rentrant en phase de reproduction sexuée ont une vitesse de sédimentation plus élevée. Selon ces auteurs, cette spécificité pourrait offrir plusieurs avantages :
Le reproduction sexuée peut être initiée chez Ditylum brightwellii par une carence en éléments nutritifs.
La densité cellulaire des populations de Ditylum brightwellii dans le milieu marin varie fortement au fil de l’année et les périodes de minima et maxima varient selon les lieux d’observation. La vitesse de reproduction asexuée de cette espèce en fait un contributeur parfois important aux blooms* algaux.
Ditylum brightwellii présente une meilleure résistance aux carences en azote qu’aux carences en phosphore, cet élément pouvant donc constituer un facteur limitant significatif. Les nitrates et l’ammonium sont stockées dans des vacuoles*, qui servent de réservoirs dans lesquels l’organisme peut puiser pour répondre à ses besoins.
Les chloroplastes* de Ditylum brightwellii présentent une mobilité marquée au sein du cytoplasme* en fonction de la luminosité. Ils se déplacent vers la périphérie des cellules dans la journée (à une vitesse de 0,1 à 0,3 µmi/mn) et rejoignent le centre de la cellule la nuit, à une vitesse plus faible. Des microtubules et des microfilaments régissent ces mouvements intracellulaires.
Ditylum brightwellii est une espèce très intéressante pour étudier la génétique des populations car elle présente d’importantes variations génétiques entre individus, y compris au sein d’une même population. De manière surprenante, la variabilité génétique reste importante dans les populations en phase de reproduction asexuée intense (bloom* algal), ce phénomène pouvant s’expliquer par une réponse adaptative aux fluctuations du milieu : les individus les plus adaptés aux conditions environnementales du moment se développent plus rapidement que les autres, mais dès que les conditions environnementales évoluent, c’est une autre sous-population, mieux adaptée aux nouvelles conditions, qui devient alors dominante. Cette variabilité génétique importante amène les scientifiques à s’interroger sur la validité de la notion d'espèce chez cet organisme. Ditylum brightwellii pourrait être en fait un consortium d'espèces qui regrouperait plusieurs espèces cryptiques* (c’est la raison pour laquelle le statut de ce taxon apparaît comme « unassessed » sur le site WORMS).
Les diatomées* sont consommées par des copépodes*. Des études ont montré que Ditylum brightwelli induit une baisse importante de la fécondité chez le copépode Calanus pacificus. Les femelles de ce copépode produisent alors des œufs anormaux qui pour plus de 90 % d'entre eux n’éclosent pas. Ce phénomène n’est pas propre à Dityllum brightwellii mais il est particulièrement marqué avec cette espèce. Cet effet serait dû à la synthèse de composés chimiques inhibiteurs, notamment des aldéhydes poly-insaturés et d’autres métabolites désignés sous le nom d’oxylipine, lorsque les diatomées sont endommagées par leurs prédateurs.
Le nom français proposé reprend l’hommage à Thomas Brightwell proposé par T. West.
Le nom de genre Ditylum provient du grec ancien [Ditylos]= double, à deux segments. Ce nom illustre la symétrie de cette espèce qui frappe lorsqu’on l’observe pour la première fois.
Le nom d’espèce a été défini en hommage à Thomas Brightwell, botaniste et phycologue britannique (1787-1868). Thomas Brightwell a décrit plusieurs diatomées courantes dans le plancton*, dont Rhizosolenia styliformis.
Numéro d'entrée WoRMS : 149023
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Heterokontophyta | Hétérokontophytes | |
| Sous-embranchement | Bacillariophytina | ||
| Classe | Bacillariophyceae | Bacillariophycées | |
| Sous-classe | Coscinodiscophycidae | ||
| Super ordre | Lithodesmiophycanae | ||
| Ordre | Lithodesmiales | ||
| Famille | Lithodesmiaceae | ||
| Genre | Ditylum | ||
| Espèce | brightwellii |
 Diatomées et autre phytoplancton
Diatomées et autre phytoplancton
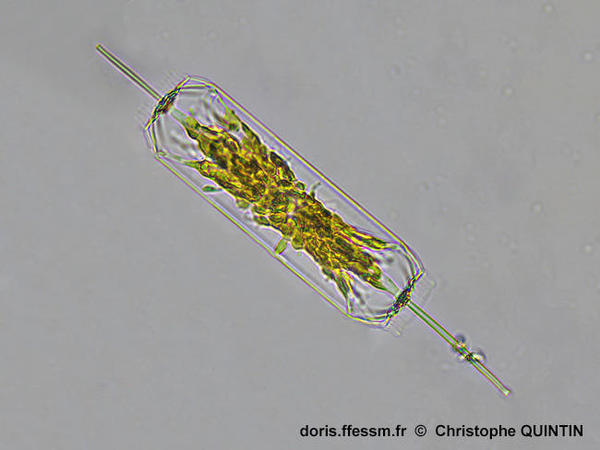
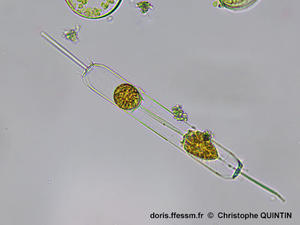
Vue d'une cellule complète en vue connective*
Cette
photographie permet de bien voir les caractéristiques essentielles de cette
espèce, avec un frustule* d'apparence rectangulaire, prolongé de chaque côté par
un processus relativement long (rimoportula*). Les extrémités du frustule comportent un velum*
constitué d'épines courtes. Les chloroplastes* sont granulaires.
Photographie
prise au microscope avec un objectif x40
Echantillon prélevé au filet à plancton (maille 35 µm) dans le port de Binic (22)
30/12/2023
Diatomées et autre phytoplancton
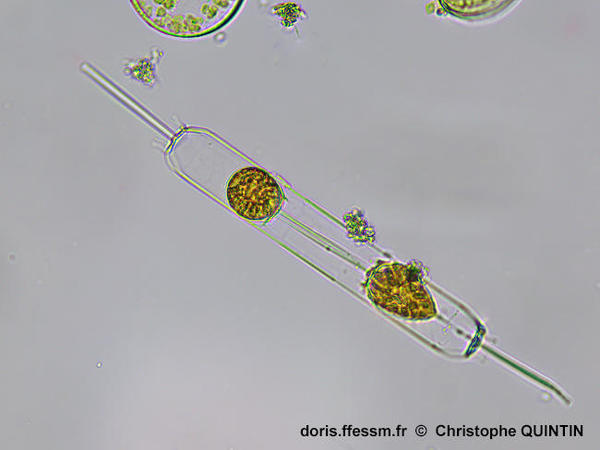
Cellule en cours de division
Lors de la
reproduction asexuée, deux individus se forment à partir d'un demi-frustule* chacun, fourni par la cellule mère.
L'un des deux individus fils sera plus petit que l'original. On est ici en fin
de processus, les deux individus étant presque totalement séparés.
Photographie prise au microscope avec un objectif x40
Echantillon prélevé au filet à plancton (maille 35 µm) dans le port de Binic (22)
18/02/2024

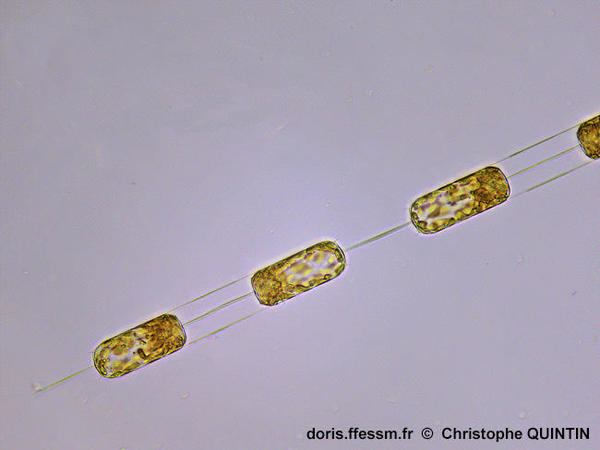
Une chaîne de quelques individus
Ditylum brightwellii peut se présenter sous la forme de courtes chaînes de quelques individus, reliés entre eux par les processus terminaux.
Photographie
prise au microscope avec un objectif x40
Echantillon
prélevé au filet à plancton (maille 35 µm) dans le port de Binic (22)
19/03/2023

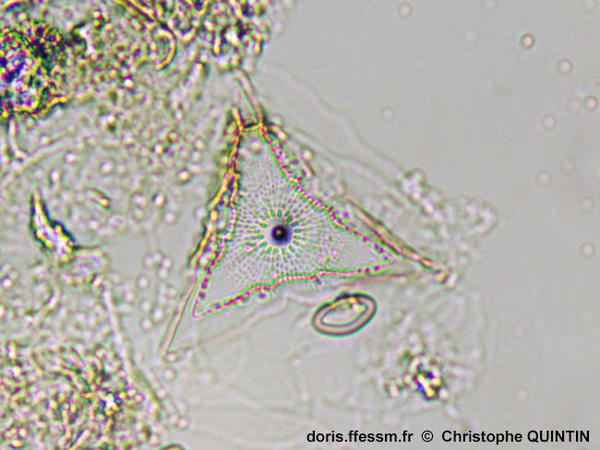
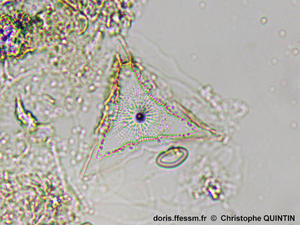
Un individu en vue valvaire*
On observe la forme triangulaire du frustule* et le débouché interne du tube central. Il y a de nombreuses aréoles* disposées radialement.
Photographie prise au microscope avec un objectif x4
Echantillon prélevé au filet à plancton (maille 35 µm) dans le port de Binic (22)
26/10/2024
Rédacteur principal : Christophe QUINTIN
Vérificateur : Jean-Luc MOUGET
Responsable régional : Yves MÜLLER
Brussaard C.P.D., Noordeloos A.A.M., Riegman R., 1997, Autolysis kinetics of the marine diatom Ditylum brightwellii (Bacillariophyceae) under nitrogen and phosphorus limitation and starvation, Journal of Phycology, 33, 980-987.
Chen S.T., Li C.W., 1991, Relationships between the Movements of Chloroplasts and Cytoskeletons in Diatoms, Botanica Marina, 34, 505-511.
Eppley R.W., Rogers J.N., 1970, Inorganic nitrogen assimilation of Ditylum brightwellii, a marine plankton diatom, Journal of phycology, 6, 344-351.
Godhe A., Rynearson T., 2017, The role of intraspecific variation in the ecological and evolutionary success of diatoms in changing environments, Philosophical transactions of the Royal Society, B 372, 20160399.
Jung S.W., Youn S.J., Shin H.H., Yun S.M., Ki J-S., Lee J.H., 2013, Effect of temperature on changes in size and morphology of the marine diatom, Ditylum brightwellii (West) Grunow (Bacillariophyceae), Estuarine, Coastal and Shelf Science, 135,128-136.
Koester J.A., Brawley S.H., Karp-Boss L., Mann D.G., 2007, Sexual reproduction in the centric marine diatom Ditylum brightwellii (Bacillariophyta), European Journal of Phycology, 42(4), 351-366.
Li C.-W., Volcani B. E., 1985, Studies on the biochemistry and fine structure of silica shell formation in diatoms, Protoplasma, 124(1-2), 10–29.
Pohnert G., 2005, Diatom/Copepod Interactions in Plankton: The Indirect Chemical Defense of Unicellular Algae, Chembiochem: a European journal of chemical biology, 6. 946-59.
Rynearson T.A., Armbrust E.V., 2000, DNA fingerprinting reveals extensive genetic diversity in a field population of the centric diatom Ditylum brightwellii, Limnology and Oceanography, 45(6), 1329–1340.
Rynearson T. A., Armbrust E. V., 2005, Maintenance of clonal diversity during a spring bloom of the centric diatom Ditylum brightwellii. Molecular Ecology, 14(6), 1631–1640.
Rynearson T.A., Newton J., Armbrust E., 2006, Spring Bloom Development, Genetic Variation, and Population Succession in the Planktonic Diatom Ditylum brightwellii, Limnology and Oceanography, 51, 1249-1261.
Uye S., 1996, Induction of reproductive failure in the planktonic copepod Calanus pacificus by diatoms, Marine Ecological Progress Series, 133, 89-97.
Waitel A., Arrison P.J., 1992, Role of sinking and ascent during sexual reproduction in the marine diatom Ditylum brightwellii, Marine Ecological Progress Series, 87, 113-122.
La page sur Ditylum brightwellii sur le site de référence de DORIS pour les algues : AlgaeBase
La page de Ditylum brightwellii sur le site de l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.



")

