

« Assemblage » entre doridien (panache branchial sur médiane dorsale) et dendronotidé (papilles dorso-latérales arborescentes)
Papilles arborescentes : 5 à 7 sur bord antérieur de la tête, 5 plus grandes sur chaque côté du dos
Epiderme jaune plus ou moins pâle et orangé, avec couverture dorsale variable de taches rouge-orangé vif
Tasselled nudibranch (GB)
Doris ramosa Cantraine, 1835
Euplocamus croceus Philippi, 1836
Idalia ramosa Philippi, 1844
Euplocamus japonicus Bergh, 1880 (1879)
Caloplocamus ramosus Pruvot-Fol, 1951
Euplocamus plumosus Schultz, in Pruvot-Fol, 1954
Méditerranée, Atlantique oriental, Afrique du Sud, océan Indien, Pacifique
Zones DORIS : ● Europe (côtes françaises), ○ [Méditerranée française], ○ [Atlantique Nord-Est, Manche et mer du Nord françaises], ● Indo-PacifiqueCette espèce est signalée en Méditerranée (Israël, Crète, Croatie, Malte, Italie, France, Espagne), en Atlantique Nord-Est (Espagne/Andalousie atlantique jusqu'au nord du golfe de Gascogne, Açores, Madère, Canaries, Selvagens, Ghana, Angola), en Atlantique SE (Afrique du Sud), dans l’océan Indien (Afrique du Sud, Kenya, Tanzanie), dans le Pacifique NW (Hong Kong, Chine/sud, Corée du Sud, Japon), dans le Pacifique N-central (Hawaii, Midway, Kure) et dans le Pacifique SW (Australie/SE, Nouvelle Zélande/NW).
Elle est apparemment absente du continent américain.
Cette espèce fréquente les fonds rocheux (dans les trous et sous les pierres) ou semi-meubles, riches en bryozoaires arborescents, de 5 à 400 m de profondeur dans des eaux tempérées chaudes à tropicales.
Kaloplocamus ramosus mesure habituellement de 30 à 35 mm de long, avec un maximum de 60 mm. Sa largeur est égale à environ 1/3 de la longueur.
Le corps, allongé, large et épais, est plus élevé entre les rhinophores* et le panache branchial*. La queue est courte, en pointe arrondie.
Le bord antérieur de la tête est bordé par un voile oral légèrement arrondi, porteur de 5 à 7 papilles arborescentes (leur nombre dépend de la taille de l’individu) entre les bases des rhinophores. Leur aspect arborescent est dû à la présence, sur leur 1/3 terminal, de plusieurs courtes papilles* secondaires digitiformes à bout arrondi.
Chaque côté du dos (bord palléal*) porte 1 rangée longitudinale de 5 papilles similaires mais plus grandes. La première papille dorsale se trouve juste en arrière et sous le niveau du rhinophore. Les suivantes se trouvent sur le côté du dos. Ces papilles dorsales n’ont aucune relation avec la glande digestive et ne peuvent donc être qualifiées de "cérate"*.
Les côtés du dos entre le rhinophore et le panache branchial sont légèrement carénés.
Les rhinophores sont rétractiles et leur tiers terminal (clavus ou ogive) est lamellé.
Le panache branchial typique des doridiens est situé sur la médiane dorsale, aux 2/3 postérieurs de la longueur du corps. Il est constitué de 5 plumes tri-pennées*, placées en fer-à-cheval autour de la papille anale. Elles peuvent se contracter (se replier) mais ne peuvent être rétractées dans une poche dorsale (Phanerobranchia).
Un tentacule oral, en forme de lamelle rectangulaire aux petits côtés arrondis, est situé en avant de chaque rhinophore, sous le voile oral (qui porte les papilles arborescentes) de part et d’autre de l’orifice buccal.
La couleur de base du corps, des troncs des papilles, des rhinophores et des branchies est jaune-safran (= jaune clair ± orangé) à orangé, parsemée de taches variables rouges à rouge-brun.
La teinte du cône du rhinophore est jaune-clair verdâtre chez les spécimens du golfe de Gascogne (cf. photos) et de Méditerranée.
Le tronc des papilles antérieures et latérales a la même teinte que l’épiderme dorsal. Les papilles secondaires sont souvent pâles à blanchâtres.
La coloration peut varier très largement, du blanc translucide jaspé d’éclaboussures jaune-orange au gris pâle moucheté de rouge-orange, au jaune-crème pâle tacheté diversement d’orange ou de brun foncé jusqu’au rouge vif presque uniforme avec un minuscule pointillé jaune-orange. Aucun des schémas chromatiques ne semble dépendant de la localité et divers motifs sont parfois observés dans la même zone.
En Europe : aucune (les Dendronotidae et les Tritoniidae possèdent des cérates arborescents mais pas de panache branchial médio-dorsal). Toutefois, en cours de plongée, en cas de faible luminosité ou de petits spécimens, la présence des papilles arborescentes peut induire une confusion avec certaines espèces de Dendronotidae (Dendronotus, Marionia blainvillea, Tritonia) ou avec Plocamopherus tilesii Bergh, 1877, espèce lesseptienne* récemment introduite dans l’est de la Méditerranée.
Aux Açores, une autre espèce de Kaloplocamus a été décrite K. (Euplocamus) atlanticus Bergh, 1899. La différence principale concerne le nombre et le type de dents de la radula*, mais il pourrait s’agir de la même espèce.
Dans le Pacifique et en Afrique du Sud, d’autres espèces de Kaloplocamus sont connues. Les caractères fournis par la littérature permettent de les différencier.
K. ramosus est un prédateur de Bryozoaires branchus (arborescents) ou qui présentent, sur leur périphérie, des tronçons assimilables à des branches comme par exemple Smittina (Porella) cervicornis. Très exceptionnellement, on pourra le trouver sur des bryozoaires encroûtants, mais sans la certitude qu’il s’en nourrisse effectivement (voir à "Divers Biologie" la Radula).
L’examen de la littérature et du contenu stomacal de 10 spécimens récoltés dans le golfe de Gascogne en 2009 et 2010 par des biologistes de l’Ifremer (campagnes EVHOE) témoigne de la consommation de Bugula neritina, Caberea boryi, Cellaria salicornioides, Scrupocellaria scabra, Smittina cervicornis, Reteporella grimaldii, Scrupocellaria cf. reptans , Scrupocellaria incurvata et Margaretta cereoides (synonyme de Tubucellaria cereoides).
On ne connait presque rien à ce sujet. La reproduction est, comme pour les autres Doridiens, sexuée et hermaphrodite*. Les organes génitaux mâle et femelle sont voisins et situés sur le flanc droit, postérieurement à la base du rhinophore. La période de reproduction n’est pas connue.
La seule information disponible sur la ponte est la photo, réalisée par Cory Pittman à Hawaii, d’un étroit ruban spiralé (diamètre 12 mm, largeur 2 mm) jaune-orangé, fixé par un côté au substrat* (voir photo).
La grande variabilité chromatique ne semble pas dépendre de la localité car divers motifs sont parfois observés dans la même zone. Cette variabilité pourrait être due, totalement ou partiellement, à des différences alimentaires ou à la présence d’espèces différentes mais de morphologie très semblable.
Une capacité de mouvement des papilles arborescentes a été décrite ainsi qu’une capacité de bioluminescence* lorsque le spécimen est importuné.
Cette espèce semble être principalement sciaphile*, avec une prédilection pour les endroits sombres (trous dans la roche, dessous de pierre, grandes profondeurs ou déplacements nocturnes), mais elle est parfois observée en plein jour, à la surface du substrat.
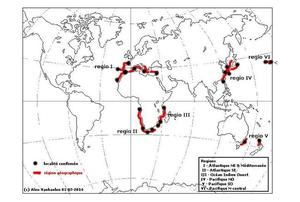
Distribution mondiale. L’espèce présente un schéma de distribution intrigant (voir photo – Distribution mondiale) : elle est signalée en Méditerranée, dans l’Atlantique NE (Madère, Açores, détroit de Gibraltar, Canaries) et SE (Angola, Ghana) en Afrique du Sud (Cape Town), l’ouest de l’océan Indien (Tanzanie, Afrique du Sud), dans le Pacifique W (Hong Kong, Corée du Sud, Chine du Sud, Japon), le Pacifique Central (Hawaii) et le Pacifique Sud (Australie SE et Nouvelle Zélande). Mais nulle part sur le continent américain.
Cela soulève quelques questions :
- s’agit-il d’une seule espèce très variable ou de plusieurs espèces proches ?
- s’il s’agit d’une seule espèce, quelle est son origine géographique ? Et quels sont les mécanismes de sa dispersion ?
- quel est son type de développement larvaire* (de celui-ci dépendent les mécanismes de sa dispersion) ?
L’analyse ADN de spécimens issus des différentes régions apportera probablement une réponse. Mais plus d’informations sur sa reproduction et sa biologie sont également nécessaires.
C’est pourquoi, toute information (biotique* et abiotique*) est la bienvenue. En cas de découverte d’un spécimen, vous pouvez contribuer à mieux connaître K. ramosus :
Je rencontre Kaloplocamus ramosus. Que puis-je faire ?
I meet Kaloplocamus ramosus. What can I do ?
Francisation du nom scientifique.
Kaloplocamus : du grec [kalos] = beau et [plokamos] = boucle de cheveux, crinière d’animal ; en allusion à l’aspect du dos orné de nombreuses papilles arborescentes.
ramosus : du latin [ramosus] = branchu, ramifié.
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Mollusca | Mollusques | Organismes non segmentés à symétrie bilatérale possédant un pied musculeux, une radula, un manteau sécrétant des formations calcaires (spicules, plaques, coquille) et délimitant une cavité ouverte sur l’extérieur contenant les branchies. |
| Classe | Gastropoda | Gastéropodes | Mollusques à tête bien distincte, le plus souvent pourvus d’une coquille dorsale d’une seule pièce, torsadée. La tête porte une ou deux paires de tentacules dorsaux et deux yeux situés à la base, ou à l’extrémité des tentacules. |
| Sous-classe | Heterobranchia | Hétérobranches | |
| Super ordre | Nudipleura | Nudipleures | |
| Ordre | Nudibranchia | Nudibranches | Cavité palléale et coquille absentes chez l’adulte. Lobes pédieux souvent absents aussi. Respiration cutanée, à l’aide de branchies, de cérates ou d’autres appendices. Tête portant une ou deux paires de tentacules, les tentacules postérieurs ou rhinophores peuvent parfois être rétractés dans des gaines. Principalement marins ou d’eau saumâtre. |
| Sous-ordre | Doridina | Doridiens | Corps aplati. Anus dorsal entouré complètement ou partiellement par des branchies de remplacement ramifiées qui peuvent être rétractées (voire absentes). Mangeurs d’éponges, habituellement armés de spicules calcaires internes. |
| Famille | Polyceridae | Polycéridés | Doridiens limaciformes aux rhinophores lamellés avec souvent quelques papilles frontales et branchiales. Présence de lobes oraux sur la tête ou de tentacules buccaux développés. |
| Sous-famille | Triophinae | Triophinés | |
| Genre | Kaloplocamus | ||
| Espèce | ramosus |
 Gastéropodes Opisthobranches
Gastéropodes Opisthobranches

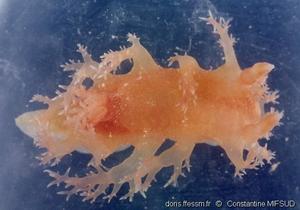
Premier spécimen observé en plongée en France
Papilles arborescentes, rhinophores et panache branchial.
Cros-de-Cagnes (06), de nuit, 20 m
25/02/2022
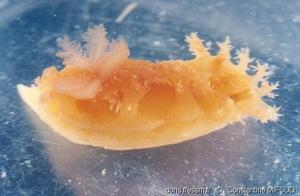
Gastéropodes Opisthobranches

Papilles arborescentes, rhinophores et panache branchial
Cros-de-Cagnes (06), 20 m, de nuit
25/02/2022


Sur le sable
Importante extension de la partie postérieure de la sole pédieuse.
Cros-de-Cagnes (06), de nuit, 20 m
25/02/2022


Vue dorsale
Les caractères utiles à l’identification sont la présence d’un panache branchial dorsal caractéristique des doridiens, associé aux deux rangées de papilles dorso-latérales qui évoquent les dendronotidés, ainsi que les papilles antérieures (habituellement 6) sur le bord de la tête (voile oral).
Un petit "col" entoure le pied des rhinophores : c’est le bord surélevé de la poche rhinophorienne.
Reggio Calabria (Italie), 12 m
20/09/2009


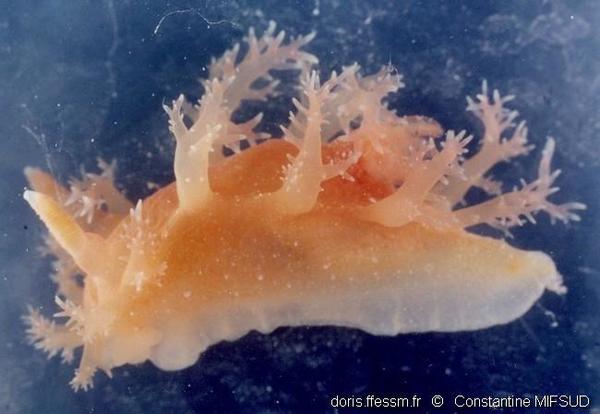
Dos et tête
Les caractères utiles à l’identification sont la présence d’un panache branchial dorsal associé à deux rangées de papilles dorso-latérales ainsi que des papilles (habituellement 6) sur le bord antérieur de la tête (voile oral).
Les lamelles sur le clavus (ogive) du rhinophore droit sont bien visibles.
Reggio Calabria (Italie), 12 m
20/09/2009


Tête
Le mucron (flèche), petite excroissance à l’apex du rhinophore droit, est bien visible, ainsi que le bord légèrement surélevé de la poche rhinophorienne.
Reggio Calabria (Italie), 12 m
02/07/2014


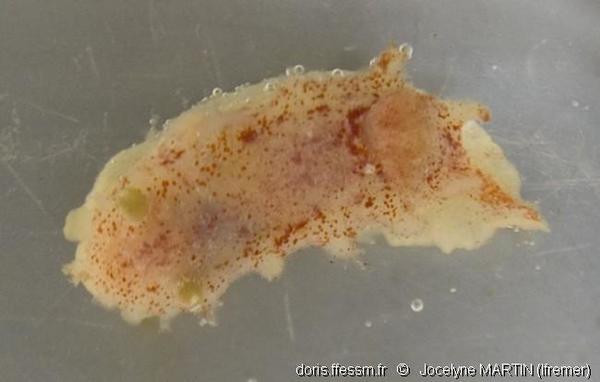
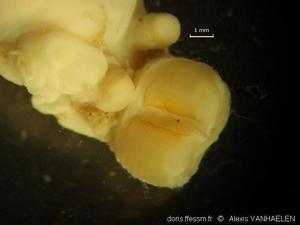
Spécimen Ifremer N0817-01 in vitro
Ce spécimen a été capturé par dragage, ce qui explique probablement les dégradations subies: absence du rhinophore gauche, lésions des papilles dorsales. Longueur ± 23 mm.
Golfe de Gascogne, 133 m (dragage)
04/11/2009

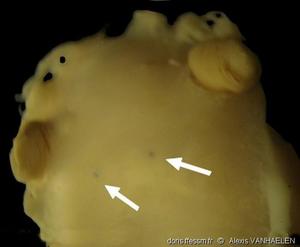
Spécimen Ifremer N0817-02 in vitro
Ce spécimen a également été capturé par dragage. Les deux rhinophores sont intacts. La tache claire à leur apex est le mucron. Longueur ± 28 mm.
Golfe de Gascogne, 133 m (dragage)
04/11/2009

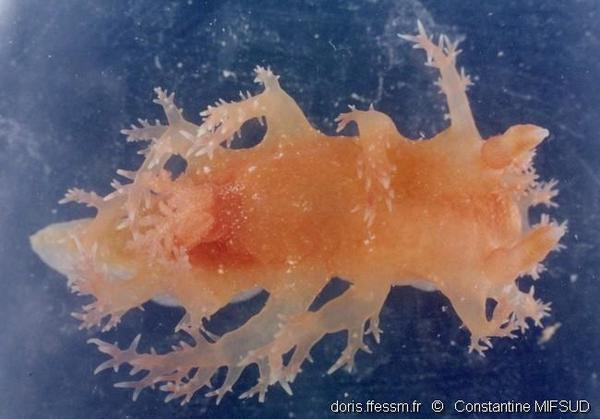
Vue dorsale
Les papilles dorsales et antérieures sont intactes et étendues. Leurs ramifications sont bien visibles, de même que le « col » à la base du rhinophore gauche. Un léger agrandissement permet de distinguer les lamelles des rhinophores et les mucrons.
Ras il-Wahx (Malte), 100 m (dragage)
05/2003

Côté gauche
Les papilles dorsales et antérieures sont intactes et étendues. Leurs ramifications sont bien visibles. Un léger agrandissement permet de distinguer les lamelles des rhinophores.
Les ponctuations blanches sont des petits dartres cutanés.
Ras il-Wahx (Malte), 100 m (dragage)
05/2003


Panache branchial
Les branchies sont tripennées.
Fomm ir-Rih (Malte), 120 m (dragage)
04/04/2005


Contracté
L’animal, partiellement contracté, forme une « bosse ».
Fomm ir-Rih (Malte), 120 m (dragage)
04/04/2005


Ponte en spirale, in vitro
Ponte déposée en aquarium par un spécimen de 17 mm, capturé en plongée de nuit ; diamètre de la spirale = 12 mm (Echelle : 10 mm).
Black Rock (Maui, Hawaii)
05/08/2001


Ponte in vitro, détail
Ponte déposée en aquarium par un spécimen de 17 mm, capturé en plongée de nuit ; diamètre de la spirale = 12 mm (Echelle : 2 mm).
Black Rock (Maui, Hawaii)
05/08/2001


Lamelles des rhinophores
Lamelles des rhinophores du spécimen n° N-0796/06. Photo prise avec une caméra SONY DSC W-120 au travers de l’oculaire d’une loupe binoculaire (grossissement x24).
Golfe de Gascogne, 142 m (dragage)
25/10/2009

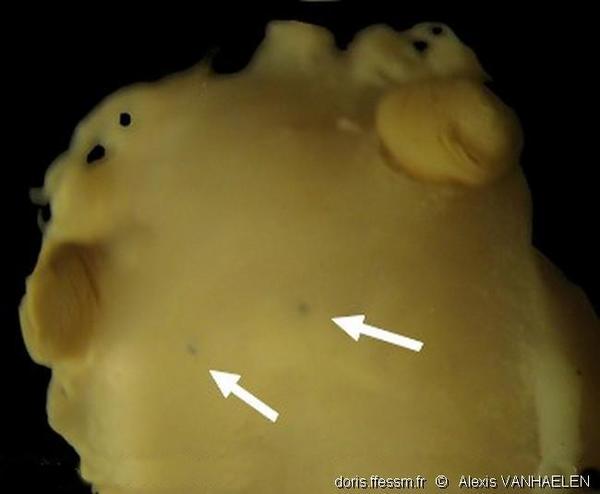
Yeux dorsaux
Yeux dorsaux du spécimen n° N-0822. Certaines espèces d’opisthobranches possèdent des yeux rudimentaires, enfouis sous l’épiderme dorsal. Les biologistes Vayssières et delle Chiaje ont montré que ces yeux sont reliés au système nerveux de l’animal. Ils semblent suffisants pour enregistrer des variations d’intensité de lumière de type "jour/nuit" ou "dans/hors d’un abri" mais pas pour avoir une vision nette de leur environnement.
Photo prise avec une caméra SONY DSC W-120 au travers de l’oculaire d’une loupe binoculaire (grossissement x32).
Golfe de Gascogne, 159 m (dragage)
05/11/2009

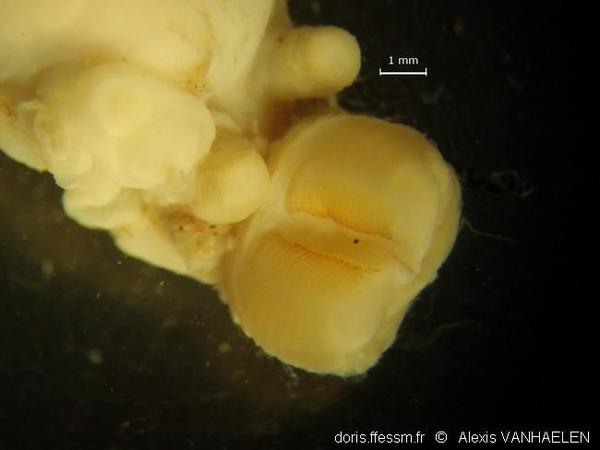
Radula
Radula du spécimen n° N-0799/07. La radula et son support (le "coussinet") sont évaginés : un artefact dû vraisemblablement au mode de collecte (dragage) ou de conservation (mise en éthanol).
Cette photo illustre bien la formule de la radula [(15-17)-3-0-3-(17-15)] :
- au centre les dents sont absentes (= 0) ;
- de chaque côté de cette zone vide, il y a 3 lignes de crochets : intacts à l’avant, ils se cassent au fil du temps (= 3) ;
- les dents des lignes suivantes (jusqu’à 17) ont une forme de tuile.
Cet assemblage permet de casser des sections de bryozoaires arborescents et de les acheminer vers le tube digestif.
Photo prise avec une caméra SONY DSC W-120 au travers de l’oculaire d’une loupe binoculaire (grossissement x24) (échelle = 1 mm).
Golfe de Gascogne, 150 m (dragage)
26/10/2009


Radula
Radula du spécimen n° N-0799/07. Cette photo, prise au microscope électronique à balayage (MEB) détaille la précédente :
- au centre les dents sont absentes (= 0) ;
- de chaque côté de cette zone vide, il y a 3 lignes de crochets (= 3) ;
- les dents des lignes suivantes (jusqu’à 25) ont une forme de tuile.
Photo prise avec le MEB de l’Institut Royal des Sciences Naturelles de Belgique, à Bruxelles, par Julien Cillis et Laetitia Despontin, auxquels nous adressons nos plus vifs remerciements.
Golfe de Gascogne, 150 m (dragage)
Direction opérationnelle Taxonomie et Phylogénie Institut Royal des Sciences Naturelles de Belgique
26/10/2009

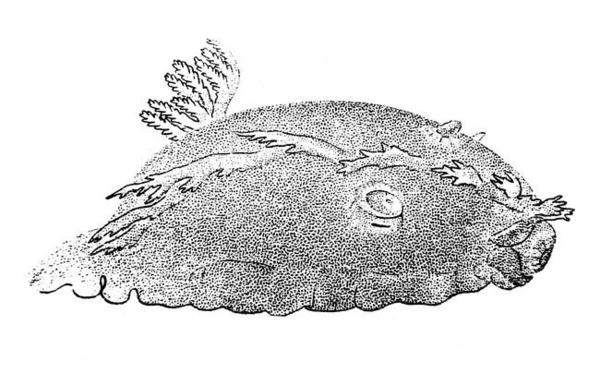
Dessin originel de Cantraine
Premier dessin de K. ramosus (en noir et blanc) sous le nom de Doris ramosa.
Cantraine F.J., 1841, MALACOLOGIE MÉDITERRANÉENNE ET LITTORALE OU DESCRIPTION DES MOLLUSQUES QUI VIVENT DANS LA MÉDITERRANÉE OU SUR LE CONTINENT DE L’ITALIE, AINSI QUE DES COQUILLES QUI SE TROUVENT SUR LES TERRAINS TERTIAIRES ITALIENS, AVEC DES OBSERVATIONS SUR LEUR ANATOMIE, LEURS MŒURS, LEUR ANALOGIE ET LEUR GISEMENT., ouvrage servant de malacologie italienne et de complément à Brocchi, CONCHIOLOGIA FOSSILE SUBAPENNINA, Vol.1., Nouveaux Mémoires de l’Académie Royale des Sciences et Belles Lettres de Bruxelles, 13: 1-175 ; pl. I-VI.
N/A
Reproduction de documents anciens
1841


Dessin en couleurs de Philippi
Premier dessin de K. ramosus en couleurs (sous le nom de Idalia ramosa).
Philippi R.A., 1844, Fauna Molluscorum viventium et in tellure tertiaria fossilium Regni Utriusque Siciliae. Enumeratio Molluscorum Siciliae cum viventium tum in tellure tertiaria fossilium quae in itinere suo observavit, Vol. 2., ed. Anton, Halle. iv, 303 pp., pl. 1-28.
N/A
Reproduction de documents anciens
1844

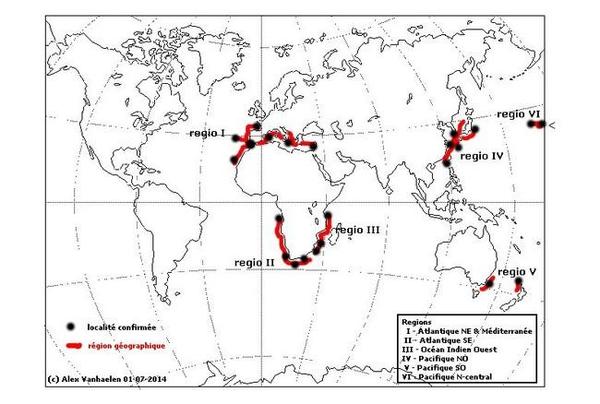
Distribution mondiale
Les zones où l’espèce a été observée sont indiquées en rouge ; les localités (ou ensembles de localités proches) sont indiquées par des points noirs.
Cette carte compile les localisations répertoriées dans la littérature et/ou affichées sur plusieurs sites Internet. Elle tient compte de toutes les synonymies connues à ce jour. Cette distribution devra être confirmée ou infirmée dans le futur par analyse ADN.
N/A
01/07/2014
Rédacteur principal : Alex VANHAELEN
Vérificateur : Jean-Pierre COROLLA
Responsable régional : Jean-Pierre COROLLA
Cantraine F.J., 1835, Diagnoses ou descriptions succinctes de quelques espèces nouvelles de mollusques, Bulletin de l’Académie Royale des Sciences et Belles Lettres de Bruxelles, 11, 380-401.
Edmunds M., 2010, Opisthobranchiate Mollusca from Ghana : Polyceridae, Journal of Conchology, 40(3), 291-303.
Vallès Y., Gosliner T.M., 2006, Shedding light onto the genera (Mollusca : Nudibranchia) Kaloplocamus and Plocamopherus with description of new species belonging to these unique bioluminescent dorids, The Veliger, 48(3), 178-205.
Vanhaelen A., Massin C., Martin J., Laffargue P., 2014, Kaloplocamus ramosus (Cantraine, 1835) (Gastropoda: Polyceridae) : new records in the Bay of Biscay, with notes on distribution and food, Iberus, 32(1), 53-64.
Zsilavecz G., 2007, NUDIBRANCHS OF THE CAPE PENINSULA AND FALSE BAY, Ed. Southern Underwater Research Group Press, Cape Town, South Africa, 104p.
La page sur Kaloplocamus ramosus dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.
















")

