

Algue unicellulaire visible au microscope optique
Squelette siliceux formé de tubes
Squelette formant un carré ou un losange
Une longue épine disposée à chaque angle du carré
Présence de 4 « fenêtres »
Présence de petites épines, orientées vers l’intérieur, sur le carré basal
Dictyocha abnormis Ehrenberg 1845
Dictyocha tenella Ehrenberg 1861
Atlantique, Méditerranée
Zones DORIS : ○ [Atlantique Nord-Est, Manche et mer du Nord françaises], ○ [Méditerranée française]Cette espèce, cosmopolite, se rencontre dans la plupart des eaux marines froides et tempérées mondiales.
En Méditerranée, l’espèce est limitée au nord-est du bassin méditerranéen, où la température de l’eau descend sous les 12 °C en hiver.
Espèce planctonique*
Dictyocha fibula est une algue unicellulaire (visible au microscope optique) qui peut se présenter sous trois formes connues : une forme nue à flagelle*, une forme nue plurinucléée et une forme dotée d’un squelette.
La forme la plus visible, faisant l’objet de cette fiche, est celle comportant un squelette siliceux formé de tubes. Ce squelette présente l’allure générale d’un carré ou d’un losange orné d’une longue épine disposée à chaque angle du carré (soit 4 épines en tout). Un « pont », fixé sur ce carré basal, s’élève au-dessus du carré pour constituer un volume dans lequel s’abrite la majeure partie de la cellule algale. Ce « pont » est arrimé de chaque côté sur deux des côtés du carré basal, ce qui conduit à la présence de 4 « fenêtres ». Il est parallèle à 2 côtés du carré basal, ou très légèrement incliné. Chaque côté du carré est orné d’une petite épine, orientée vers l’intérieur. Les épines externes peuvent être de longueur similaire mais il arrive que les paires constituées des épines opposées aient des longueurs différentes l’une de l’autre (voir photo). Une épine apicale*, fixée au « pont » et perpendiculaire au plan du carré basal, donc difficile à voir en microscopie, est en général présente (absente de la forme messanensis notamment).
Cette algue, encore plus que la plupart des silicoflagellés, présente d’importantes variations morphologiques intraspécifiques, mais les éléments évoqués ci-dessus sont généralement présents, même s’ils peuvent disparaître chez des formes régressées. Il existe une forme, dite hexagona présentant 6 branches, ce qui pourrait permettre la confusion avec Octactis speculum, mais l’absence d’anneau distal dissipe rapidement une éventuelle confusion.
Le diamètre global du squelette peut atteindre 90 microns, le diamètre de l’anneau basal étant de 45 microns.
Chez les algues vivantes, le protoplasme* (le contenu de la cellule) occupe la quasi-totalité de l'espace circonscrit par le squelette, et il donne naissance à de nombreux prolongements ou pseudopodes* très fins. D'après la littérature, le positionnement exact de ce squelette n'est pas clair. En effet, Chang et al.(2017) observent que les éléments du squelette sont positionnés à l'extérieur de la cellule (images prises en microscopie électronique à transmission), mais ils indiquent que ce squelette est interne dans la diagnose de l'espèce... La confusion n'est peut-être qu'apparente car dans une autre étude, il a été observé, chez une autre espèce de Dictyocha (D. octonaria), que lors de la division cellulaire (ou mitose), les éléments du squelette sont mis en place dans la jeune cellule, avant d'être externalisés lorsque la cellule atteint sa forme et sa taille définitive (Chang, 2015).
La cellule algale comporte un pyrénoïde* au sein de chaque chloroplaste*, ces derniers, petits et discoïdes, de couleur jaunâtre à brun-verdâtre, étant plutôt positionnés en périphérie et très nombreux. Elle contiendrait près de 70 appareils de Golgi (ces derniers régulent le transport et la modification des protéines synthétisées, seule une quinzaine sont visibles simultanément en microscopie électronique à transmission). Le noyau est situé au centre de la cellule et est séparé des chloroplastes périphériques par des vacuoles* (caractéristique retrouvée chez toutes les dictyochales).
Sur les spécimens vivants, il est possible d’observer la présence d’un long flagelle*, courant le long d’une des épines. Ce flagelle est antérieur (donc tirant la cellule lors de ses déplacements). En microscopie électronique, il est possible d’observer la présence d’un second flagelle, limité à sa partie basale et donc très court (0,6 microns).
En culture expérimentale, Van Valkenburg et Norris (1970) ont observé que les cellules nues venaient parfois se fixer au fond du récipient de culture et abandonnaient alors leur flagelle et toute mobilité, continuant à se reproduire de manière asexuée. Ces cellules se divisent parfois sans cytokinèse*, conduisant ainsi à l’apparition de cellules plurinuclées*.
Le silicoflagellé Dictyocha crux possède également quatre épines et peut donc être confondu avec Dictyocha fibula. On l’en distingue cependant assez facilement car Dictyocha crux possède une cinquième fenêtre, centrale.
Dictyocha fibula est une algue autotrophe* photosynthétique*. L'algue tire son énergie de la lumière solaire, et grâce à la dissociation des molécules d'eau et à l'absorption de dioxyde de carbone et des sels minéraux dissous dans l’eau, elle fabrique les matières organiques nécessaires à son développement.
Comme la plupart des silicoflagellés, Dictyocha fibula utilise abondamment la reproduction asexuée, par division. La reproduction asexuée de la forme à squelette conduit à l’existence d’individus à double squelette (individus en cours de division). Le second squelette est construit en miroir au premier et positionné face au carré basal. Les deux squelettes sont très proches l’un de l’autre, au point qu’ils sont initialement indissociables. Contrairement à ce qui se produit avec les diatomées*, la taille du squelette fils est similaire à celle du squelette père. La reproduction asexuée maintient donc la taille des individus dans la colonie.
Comme chez Octactis speculum, la manière dont s’agencent les différentes formes de Dictyocha fibula dans le cycle de vie de cette espèce n’est pas encore connue.
Il n’y a pas de reproduction sexuée connue chez cette espèce aujourd’hui.
L’optimum thermique de cette espèce se situe entre 10 °C et 12 °C., alors qu’elle a longtemps été considérée, à tort, comme une espèce préférant les eaux chaudes. Van Valkenburg et Norris (1970) ont ainsi cultivé l’espèce, qui montrait un optimum de croissance à 10 °C pour une concentration en sel de 24 ‰. En Méditerranée, l’espèce se développe principalement en hiver et au début du printemps (jusqu’à fin mars). Lorsque les eaux se réchauffent, dès qu’elles dépassent 15 °C, les individus sont rencontrés en eau plus profonde (au-delà de 50 m). Cette préférence pour la croissance hivernale se rencontre également dans des secteurs plus septentrionaux.
Les chloroplastes* de Dictyocha fibula contiennent de la chlorophylle* a et c ainsi que de la fucoxanthine.
L’étude au microscope électronique d’individus cultivés met en évidence de très petites épines à l’extrémité de chaque barreau. Ces épines ressemblent à celles rencontrées à l’extrémité des spicules* des hexactinellides, ce qui pourrait laisser penser à un mode de fabrication similaire. De telles petites épines sont observables au point de jonction entre les barreaux, ce qui laisse penser que la fabrication du squelette de Dictyocha fibula par la cellule est séquentielle, barreau par barreau, avec une soudure finale des barreaux sur un barreau préexistant afin de construire cette structure en réseau caractéristique.
Dictyocha fibula est réputée pour être sensible aux intrants, son développement en milieu marin étant favorisé par les apports d’eau douce.
Le naturaliste prussien Christian Gotfried Ehrenberg (1795-1876) a décrit initialement cette espèce (en 1839) à partir de l’observation d’un squelette fossile en provenance d’un étage Miocène (de - 23 à - 5 millions d'années) d’Oran (Algérie). Plus tard, il a été constaté que cette espèce était toujours présente dans nos eaux. De ce fait, le nom proposé par Ehrenberg n’aurait pas du être retenu pour les espèces vivantes, en application des règles du code international de nomenclature (article 1.2), tout comme le nom des taxons supérieurs qui en découlent (dictyochales, dictyochaceae, dictyochophyceae).
Les silicoflagellés, groupe auquel appartient Dictyocha fibula, sont aujourd’hui représentés par un petit nombre d’espèces vivantes. Ces algues ont eu une importance considérable dans certaines périodes géologiques et l’accumulation de leurs squelettes siliceux, avec ceux de diatomées*, a conduit à la création de roches siliceuses particulières (les diatomites). Compte tenu de leurs préférences bien spécifiques en matière de température, les silicoflagellés sont fréquemment utilisés en paléoclimatologie pour évaluer la température des océans (notamment en évaluant les ratios entre différentes espèces). Ils peuvent également être utilisés pour étudier les évolutions de l’apparition du phénomène El Niño dans le temps.
La classification des silicoflagellés a très longtemps été fondée sur l’étude morphologique de leur squelette siliceux (principalement celui des espèces fossiles), induisant des évolutions importantes selon les auteurs. La taxonomie des Dictyochales, ordre auquel appartient Dictyocha fibula, a fait l’objet d’une profonde révision très récemment (2017), sur la base d’une étude fondée sur des données morphologiques, ultrastructurales, biochimiques et moléculaires.
Silicoflagellé à 4 branches : c'est le nom le plus explicite. Certains noms scientifiques sont peu attractifs pour les franciser.
Dictyocha : le préfixe dictyo- provient du grec ancien [dictyon ]= filet, treillis. Le suffixe [-cha] provient également du grec ancien [ocha] signifiant « support, structure ». Le nom de genre signifie donc littéralement « structure en forme de filet ».
fibula : provient directement du latin [fibula] = broche pour vêtement ou cheveux, l’allure générale de l’espèce ressemblant aux têtes de broches décoratives.
Numéro d'entrée WoRMS : 157463
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Ochrophyta | Ochrophytes | ou Hétérokontes, ou Straménopiles: présence d'un stade unicellulaire à 2 flagelles, un lisse et un à poils tubulaires. |
| Classe | Dictyochophyceae | Dictyochophycées | |
| Ordre | Dictyochales | ||
| Famille | Dictyochaceae | Dictyochacées | |
| Genre | Dictyocha | ||
| Espèce | fibula |
 Diatomées et autre phytoplancton
Diatomées et autre phytoplancton

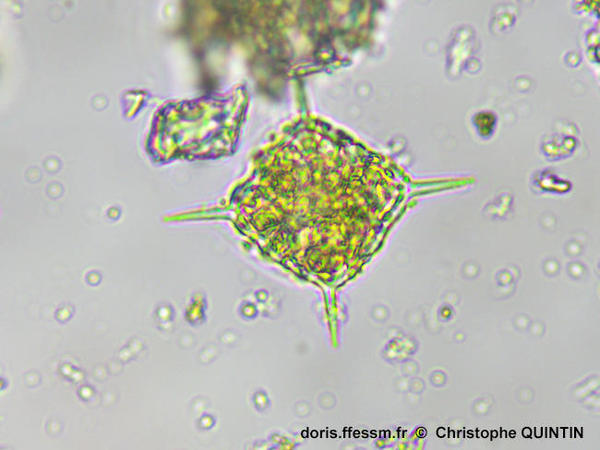
Vue au microscope d'un squelette
On peut observer les quatre épines et les quatre barreaux de la base donnant l'allure générale au squelette ainsi que le "pont" venant s'appuyer sur les barreaux.
Photographie au microscope d'un individu prélevé dans le port de Binic (22), objectif x 40, image obtenue par empilement de plusieurs images afin d'obtenir une image nette sur la totalité de la hauteur du spécimen.
26/10/2024
Diatomées et autre phytoplancton
Un individu vivant
La forme du
squelette reste visible, le protoplasme, de couleur dorée, entoure les éléments
du squelette.
Photographie
au microscope d'un individu prélevé dans le port de Binic (22), objectif x 40.
04/10/25025
Rédacteur principal : Christophe QUINTIN
Vérificateur : Jean-Luc MOUGET
Responsable régional : Yves MÜLLER
Ajuzie C., Houvenaghel G., 2015, A first record of extant silicoflagellates in coastal waters of Nigeria, Nature and Science,13(3).
Chang F.H., 2015, Cell morphology and life history of Dictyocha octonaria (Dictyochophyceae, Ochrophyta) from Wellington Harbour, New Zealand, Phycological Research, 63, 253-264.
Chang F. H., Sutherland J., Bradford-Grieve J., 2017, Taxonomic revision of Dictyochales (Dictyochophyceae) based on morphological, ultrastructural, biochemical and molecular data, Phycological Research, 65 (3), 235–247.
Ehrenberg C. G., 1839, Über die Bildung der Kreidefelsen und des Kreidemergels durch unsichtbare Organismen, Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, 1838, 59-147, pls I-IV, tables 1, 2.
Hargrave P.E., 2022, Actual and Potential Harmful Microalgae from the Indian River Lagoon System, Florida, USA, Technical Report #200, Harbor Branch Oceanographic Institute at Florida Atlantic University
Hernández-Becerril D., Bravo-Sierra E., 2001, Planktonic Silicoflagellates (Dictyochophyceae) from the Mexican Pacific Ocean, Botanica Marina, 44, 417-423.
Meunier A., 1919, Microplankton de la Mer Flamande: 4. Les Tintinnides et cœtera, Mémoires du Musée Royal d'Histoire Naturelle de Belgique = Verhandelingen van het Koninklijk Natuurhistorisch Museum van België, VIII(2), Hayez, imprimeur de l'Académie royale de Belgique, Bruxelles, 59, 2 plates pp.
Nival P., 1965, Sur le cycle de Dictyocha fibula Ehr. dans les eaux de surface de la rade de Villefranche-sur-mer, Cahiers de Biologie Marine, 6(1), 67-82.
O’Kane J.A., 1970, Silicoflagellates of Monterey Bay, California, thèse de Master of Science, Faculty of the Department of Geology, San Jose State College.
Rigual-Hernandez A., Bárcena M., Sierro F., Flores J., Hernández-Almeida I., Sanchez-Vidal A., Palanques A., Heussner S., 2010, Seasonal to interannual variability and geographic distribution of the silicoflagellate fluxes in the Western Mediterranean, Marine Micropaleontology, 77, 46–57.
Sournia A., Belin C., Berland B., Erard-Le Denn E., Gentien P., Grzebyk D., Marcaillou-Le Baut C., Lassus P.., Partensky F. , 1991, Le phytoplancton nuisible des côtes de France - De la biologie à la prévention, IFREMER, Rapport 4210.
Travers A., Travers M., 1968, Les Silicoflagellés du Golfe de Marseille, Marine Biology, 1(4), 285–288.
Van Valkenburg S.E., Norris R., 1970, The growth and morphology of the silicoflagellate Dictyocha fibula Ehrenberg in culture, Journal of Phycology, 6, 48 - 54.
Van Valkenburg S.E., 1971, Observations on the fine structure of Dictyocha fibula Ehrenberg, I. The skeleton, Journal of Phycology, 7(2), 113–118.
Valkenburg S.E., 1971, Observations on the fine structure of Dictyocha fibula Ehrenberg, II. The protoplast, Journal of Phycology, 7(2), 118–132.
La page sur Dictyocha fibula sur le site de référence de DORIS pour les algues : AlgaeBase
La page de Dictyocha fibula dans l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.


")

