

Crustacé
filiforme d’apparence squelettique
Appendices
allongés, longues antennes robustes
Petites épines visibles sur les principaux
segments du
corps
Mâles plus grands que les femelles, avec
segments plus longs et griffes plus grandes
Corps
souvent tacheté de rouge chez les adultes
Attaché
au substrat par les appendices postérieurs
Souvent en grands groupes
Caprelle squelettique japonaise.
Le mot "chevrolle" a été très utilisé au XIXème siècle pour désigner différentes espèces de caprelles.
Pour désigner cette espèce-ci, DORIS conserve le nom retenu par l'INPN : caprelle japonaise.
Japanese skeleton shrimp (GB), Asiatische Gespensterkrebse (D), Harig spookkreeftje, Machospookkreeft (NL), Japansk spøkelseskreps (Norvégien)
Caprella macho Platvoet, de Bruyne & Gmelig Meyling, 1995
Caprella muticum (erreur occasionnelle)
Mer du Japon, Atlantique Nord, mer du Nord, Pacifique
Zones DORIS : ● Indo-Pacifique, ● Atlantique Nord-Ouest, ● Europe (côtes françaises), ○ [Atlantique Nord-Est, Manche et mer du Nord françaises]Caprella
mutica a été décrite comme espèce indigène par Schurin en 1935 en Russie (golfe de Pierre le Grand, mer du Japon). Cependant, elle est considérée comme une des espèces invasives dont la propagation est la plus rapide en Europe. Ainsi, ces cinquante dernières années, C. mutica a été trouvée sur trois continents en sus de son continent d’origine.
La dispersion primaire de C. mutica serait liée au transport d’organismes pour l’aquaculture, aux salissures sur les coques des navires et aux eaux de ballast.
La dispersion secondaire de l’espèce le long des côtes pourrait être assurée par la navigation commerciale et de plaisance, la dérive des individus sur des macroalgues et des débris flottants.
Les populations de C.mutica associées aux zones d'intenses activités humaines sont généralement denses..
Les
caprelles colonisent volontiers des structures filamenteuses ou
arborescentes telles les algues, les hydrozoaires et les bryozoaires.
Dans son habitat d'origine, la mer du Japon, C.
mutica est présente dans l'espace
infralittoral* entre 0,7 m et 13 m de profondeur, dans des herbiers de
zostères marines ou d’algues et sur les macroalgues dérivantes, comme Sargassum muticum.
Cette algue, qui peut coloniser
l'huître japonaise Magallana gigas, est également invasive. La conchyliculture est ainsi soupçonnée
d'être un vecteur de dispersion de C.
mutica.
Dans ses eaux d’adoption de l’hémisphère nord, C.
mutica est surtout, voire uniquement,
retrouvée sur des structures artificielles comme des filets,
cordages et cages utilisés en aquaculture, des pontons de marinas et
de ports, des plateformes de forage pétrolier et des bouées de
navigation. Les substrats* artificiels pourraient constituer un
habitat transitoire où C. mutica
s’acclimaterait quelques années avant de se propager dans les
habitats naturels de son nouvel environnement.
La
tolérance de C. mutica
à une gamme étendue de températures et de salinités en fait une
candidate idéale pour l’introduction et la survie aux latitudes
tempérées à subarctiques, où seuls les milieux saumâtres (par
exemple, les têtes d’estuaires) lui seraient inhospitaliers.
La longueur totale des caprelles est mesurée de l’extrémité antérieure de la tête jusqu’à l’extrémité postérieure de l’abdomen*. Les mâles matures de C. mutica peuvent approcher ou dépasser 50 mm de longueur totale. Les femelles seront environ deux fois plus petites.
Le corps peut être divisé en trois parties : la tête (céphalon*), le thorax (péréion*) et l'abdomen.
La tête est sphérique ou ovale et dépourvue d’épines ou tubercules. Le thorax est divisé en sept segments et représente la majeure partie de la longueur du corps. La tête est soudée au premier segment thoracique tandis que l'abdomen, très petit, prolonge l'extrémité du septième segment.
Chez les mâles, les deux premiers segments thoraciques sont allongés, le deuxième étant le plus long de tous. La présence de longues soies* leur donne une apparence poilue. Le deuxième segment porte deux à trois paires d'épines sur le dos, avec deux paires additionnelles sur les côtés près de la base des membres. Les segments restants (troisième à septième) ne portent pas de soies denses mais sont pourvues d'épines tant dorsales que ventrales.
Les femelles diffèrent des mâles par des segments thoraciques beaucoup plus courts plutôt dépourvus de soies. L'ensemble tête-premier segment thoracique peut posséder une seule paire d'épines.
C. mutica possède deux paires d'antennes*, la première paire représentant plus de la moitié de la longueur totale du corps. Les segments des pédoncules* (base) sont trois fois plus longs que les flagelles ("fouets" aux extrémités des antennes). La deuxième paire d'antennes est inférieure à la moitié de la longueur de la première paire. Elles possèdent deux rangées de longues soies sur les faces ventrales des segments de leur pédoncule.
Les mandibules* et les maxillules* sont présentes à la face antérieure ventrale de la tête. Les maxillipèdes*, une paire modifiée d'appendices, servent également de pièces buccales accessoires.
Les appendices qui proviennent des segments thoraciques sont des péréiopodes* (pattes-mâchoires ou marcheuses).
Les deux premières paires, appendices de préhension, sont appelés gnathopodes*. Ils ressemblent un peu aux pattes ravisseuses des mantes-religieuses. La deuxième paire de gnathopodes est très grande et porte deux grandes épines.
Les troisième et quatrième péréiopodes sont absents et remplacés par deux paires de branchies* ovales allongées
Chez les femelles matures, deux poches marsupiales (utiles pour la reproduction) se développent également au niveau de ces segments. Ces poches sont formées par des oostégites* (expansions en forme de plaques à partir des segments basaux des appendices).
Les appendices V à VII, de longueur croissante, sont utilisés par l'animal pour s'accrocher à son substrat.
Les caprelles subissent d’importants changements de forme au cours de leur développement et de leur maturation. Cela rend leur taxonomie très complexe et il n’est donc pas rare que les jeunes et les adultes d’une même espèce soient attribués à deux espèces différentes. Ainsi, de manière générale, il est très difficile d'identifier les caprelles à l'œil nu.
C. mutica est omnivore opportuniste et consomme essentiellement les organismes associés à son substrat parmi lesquels des diatomées (végétaux unicellulaires microscopiques), des petits crustacés de type copépodes et des détritus.
Ces caprelles peuvent être brouteuses, filtreuses*, racleuses ou détritivore*-nécrophage* selon les circonstances ou le stade de développement.
En captivité, C. mutica pratique volontiers la nécrophagie cannibale ou même la prédation cannibale. Sa flexibilité alimentaire, tant en choix de proies qu'en mode d'alimentation, lui confère une grande adaptabilité et favorise sa capacité invasive.
Comme
tous les crustacés, les femelles ne sont capables de s'accoupler que
peu de temps après la mue*.
Les mâles touchent de façon répétée
les exosquelettes* des femelles avec leurs antennes pour détecter les
signes de mue (exuviation*). L'accouplement dure de 10 à
15 minutes. Une fois accouplés, les mâles défendent les femelles
pendant une courte période (environ 15 minutes). Après cette
période, les femelles montrent un comportement agressif
et chassent les mâles.
Elles vont ensuite aligner leurs orifices
génitaux (situés sur le cinquième segment thoracique) avec l'ouverture
des poches marsupiales pour y déposer des œufs fécondés. Leur nombre peut varier entre 15 et 363 par femelle (74 œufs en moyenne) chez C. mutica et il existe une corrélation positive entre la fécondité et la longueur des femelles.
L’incubation des œufs dure de 30 à 40 jours à des températures
variant entre 13 et 17 ºC et 60 jours à une température variant
entre 0 et 2 ºC. Il peut y avoir deux périodes annuelles de reproduction, l’une au
printemps, de mars à mai, et l’autre à l’été, de juin à
juillet, donnant lieu à deux générations par années.
Comme tous les amphipodes, les caprelles n'ont pas de stade larvaire* planctonique*. Le développement est dit "direct" et les nouveau-nés ressemblent à des adultes miniatures.
Les juvéniles mesurent environ 1,3 à
1,8 mm et croissent en moyenne de 0,4 à 0,9 mm par stade de mues
successives (une fois par semaine). C. mutica devient sexuellement différenciée au cinquième stade de développement. Les femelles peuvent muer plusieurs fois à l'âge adulte, devenant sexuellement réceptives à chaque mue jusqu'à la mort.
Le dimorphisme sexuel de taille et de forme est très net chez C. mutica !
Les mâles se battent en présence de femelles
réceptives à l'accouplement. Ces rencontres se terminent souvent
par la mort d'un des protagonistes car la deuxième paire d'appendices, munie de dents
empoisonnées, peut être utilisée pour empaler ou trancher un
adversaire en deux.
Cette concurrence conduisant à une forte
mortalité masculine pourrait expliquer pourquoi les populations
sauvages de C. mutica
montrent un nombre plus élevé de femelles que de mâles. En sus,
leur taille plus grande rend les mâles plus visibles et donc plus
vulnérables vis-à-vis des prédateurs.
C.
mutica et les caprelles en général
sont de piètres nageurs et peuvent passer leur vie entière
agrippées au substrat en position dressée.
Dans son habitat
natif, C. mutica
sera communément retrouvée sur des touffes fixées ou flottantes de
macroalgues de type Sargassum muticum,
S. miyabei,
S. pallidum,
Neorhodomela larix,
Polysiphonia morrowii,
Cystoseira crassipes,
Laminaria japonica,
Chrondrus
spp. et Desmarestia viridis.
Elle apprécie aussi les plantes marines telles que les zostères ainsi que
les hydrozoaires ou les bryozoaires.
Dans ses habitats d'introduction, elle tend à rechercher des organismes qui lui permettront de se camoufler. Cela inclut des macroalgues comme Ulva lactuca, Ceramium spp., Plocamium spp., Cladophora spp., Chorda filum, Fucus vesiculosus, Pylaiella spp. ou encore des hydrozoaires comme Obelia spp. et Tubularia indivisa, des bryozoaires, des amphipodes constructeurs de tubes comme Monocorophium acherusicum et Jassa marmorata. On la retrouve aussi sur des tuniciers à corps mou comme Ascidiella aspersa et Ciona intestinalis.
La durée de vie moyenne de C. mutica en laboratoire est de 68,8 jours pour les mâles et de 82 jours pour les femelles.
La
fréquente association entre certaines espèces de caprelles et des
hydrozoaires a mené à l’hypothèse d’une relation de
commensalisme*, les caprelles se
nourrissant à même les particules ou organismes captés par les
hydrozoaires ou même de mutualisme* par lequel les hydrozoaires
bénéficient d’une certaine protection offerte par les caprelles
contre les nudibranches éolidiens.
C.
mutica est
riche en acides gras polyinsaturés et en caroténoïdes*. Sa relative
abondance et son taux de croissance rapide en font une proie de choix
pour de nombreux organismes comme les poissons ou les crabes. Parmi
les autres prédateurs possibles, on pourra citer des nudibranches,
des étoiles de mer, des vers némertes, des anémones et des
hydrozoaires.
C. mutica a une très forte tolérance vis-à-vis des contraintes environnementales. Elle possède une croissance rapide et un taux de reproduction élevé. Son caractère agressif ainsi qu'une diète flexible et un fort commensalisme avec les activités humaines en font un excellent envahisseur.
Caprelle japonaise : traduction du nom anglais, qui met en avant son origine géographique.
Caprella
: du latin [capra] = chèvre ou
petite chèvre. Diminutif.
Les mouvements de la caprelle pour retrouver un
support quand elle en a été détachée pourraient évoquer des
sauts de chèvre ou de cabri (supposition des rédacteurs).
mutica
: du latin [muticus] = qui a subi une mutilation, peut
s'interpréter comme émoussé, épointé.
| Termes scientifiques | Termes en français | Descriptif | |
|---|---|---|---|
| Embranchement | Arthropoda | Arthropodes | Animaux invertébrés au corps segmenté, articulé, pourvu d’appendices articulés, et couvert d’une cuticule rigide constituant leur exosquelette. |
| Sous-embranchement | Crustacea | Crustacés | Arthropodes à exosquelette chitineux, souvent imprégné de carbonate de calcium, ayant deux paires d'antennes. |
| Classe | Malacostraca | Malacostracés | 8 segments thoraciques, 6 segments abdominaux. Appendices présents sur le thorax et l’abdomen. |
| Sous-classe | Eumalacostraca | Eumalacostracés | Présence d’une carapace recouvrant la tête et tout ou partie du thorax. |
| Super ordre | Peracarida | Péracarides | Les femelles sont dotées d'une cavité d'incubation formée par des expansions lamelleuses des péréiopodes. |
| Ordre | Amphipoda | Amphipodes | Péracarides comprimés latéralement, dépourvus de carapace, et possédant de nombreuses paires d'appendices souvent modifiés. Ils sont représentés par les gammares, les talitres, les caprelles... |
| Sous-ordre | Senticaudata | ||
| Famille | Caprellidae | Caprellidés | |
| Genre | Caprella | ||
| Espèce | mutica |
") Crustacés Malacostracés (crabes, crevettes...)
Crustacés Malacostracés (crabes, crevettes...)

Distribution : en baie de Seine
Caprella mutica caractérisée par ses antennes robustes et la présence de nombreuses soies.
Epave du Northgate, Baie de Seine (49° 30' 17.52", -00° 14' 5.58"), oc"an Atlantique, entre 20 et 25 m
23/04/2017
Crustacés Malacostracés (crabes, crevettes...)

Regard
La tête est sphérique ou ovale et elle est dépourvue d’épines ou de tubercules;
Zélande, Pays-Bas, mer du Nord
07/07/2019


Extrémité du gnathopode 2
Au niveau des pattes, les deux premières paires d'appendices sont destinés à la préhension. Ils sont appelés gnathopodes et ressemblent un peu aux pattes ravisseuses des mantes-religieuses.
Voici l'extrémité du second gnathopode. De grande taille, il porte deux grandes épines.
Zélande, Pays-Bas, mer du Nord
07/07/2019


Poils et épines
La présence de longues soies sur les segments thoraciques leur donne une apparence poilue. Le deuxième segment porte deux à trois paires d'épines sur le dos, avec deux paires additionnelles sur les côtés près de la base des membres.
Zélande, Pays-Bas, mer du Nord
07/07/2019

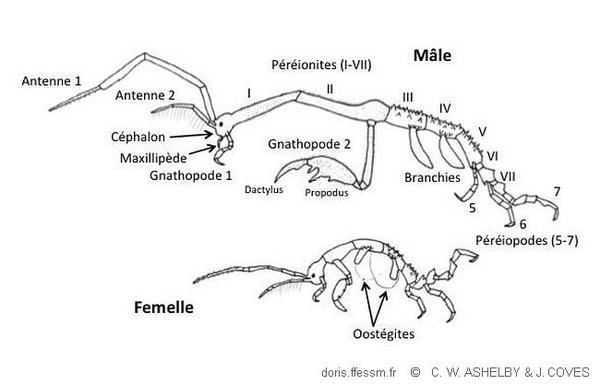
Illustration du dimorphisme sexuel
Les mâles matures de C. mutica peuvent approcher ou dépasser 50 mm de longueur totale. Les femelles seront environ deux fois plus petites..
Dessin muet présenté par [Tucotte et Sainte Marie 2009] et tiré d'[Ashelby 2005] avec la permission de l'auteur.
Annoté par le rédacteur de la
fiche.
Reproduction de documents anciens
2005


Femelle avec œufs
Après reproduction, les femelles vont aligner leurs orifices génitaux (situés sur le cinquième segment thoracique) avec l'ouverture des poches marsupiales pour y déposer des œufs fécondés.
Zélande, Pays-Bas, mer du Nord
07/07/2019


Sur un filin de bouée
Population dense de C. mutica sur un filin de bouée colonisé par des moules. Les femelles se distinguent facilement par leur poche marsupiale pleine d'œufs embryonnés. Les antennes pourvues de soies sont également parfaitement visibles.
Port de Scharendijke, Zélande, Pays-Bas, 8 m
07/2014


Détail de la tête
Détail de la tête avec le départ des deux paires d'antennes et la première paire de pattes thoraciques situées sous les maxillipèdes.
Echantillonnage dans le Ria d'Etel, Bretagne Sud, photo prise sous loupe binoculaire
24/02/2011


Deuxième paire de pattes thoraciques
Les gros appendices antérieurs sont des organes de préhension mais ils peuvent être des armes fatales quand les mâles se battent pour l'accès aux femelles réceptives à l'accouplement. Leurs dents empoisonnées peuvent être utilisées pour empaler ou trancher un adversaire en deux.
Echantillonnage dans le Ria d'Etel, Bretagne Sud, photo prise sous loupe binoculaire
02/05/2011


Femelle accrochée
Cette femelle accrochée sur un hydraire exhibe sa poche incubatrice qui se développe dans les 3ème et 4ème segments thoraciques. On distingue la paire de branchies du 3ème segment.
Echantillonnage dans le Ria d'Etel, Bretagne Sud, photo prise sous loupe binoculaire
19/10/2008


Espèce invasive
Caprella mutica est une espèce invasive qui se développe et se propage rapidement. Des densités très importantes d'individus sont observées sur des supports artificiels tels que cordages (cette photo), bouées, cages immergées, pontons.
Port de Scharendijke, Pays-Bas, 8 m
07/2014


Distribution : en Zélande
Femelle zélandaise, devant une synascidie.
La Zélande (en néerlandais : Zeeland, soit littéralement « Terre de la mer ») est une province maritime du sud-ouest des Pays-Bas. Composée d'îles et de presqu'îles, la Zélande est bordée à l'ouest par la mer du Nord (notre photo), au sud par la frontière belge, à l'est par le Brabant-Septentrional et au nord par la Hollande-Méridionale.
Zélande, Pays-Bas, mer du Nord
07/07/2019
Rédacteur principal : Equipe DORIS
Rédacteur : Jacques COVES
Correcteur : Pierre NOËL
Responsable historique : Sylvie DIDIERLAURENT
Responsable régional : Alain-Pierre SITTLER
Arimoto I., 1976, Taxonomic studies of caprellids (Crustacea,
Amphipoda, Caprellidae) found in the Japanese and adjacent waters, Spec. Publ. Seto Mar. Biol. Lab., Ser. III, 229 p.
Ashelby C., 2005, The occurrence and distribution of non-native fauna in Harwich Harbour andthe Stour and Orwell estuaries, including new records of Caprella mutica Schurin 1935 and Bugula stolonifera Ryland 1960, Essex Natur., 22, 103–116.
Boos
K., Ashton G. V., Cook E. J., 2011,
The Japanese Skeleton Shrimp
Caprella mutica (Crustacea, Amphipoda): A Global Invader of Coastal
Waters, in “In the wrong place : alien marine crustaceans -
distribution, biology and impacts”, B. Galil and P. Clark ed.,
Springer-Verlag, Berlin, Heidelberg, series in
Invation Ecology,
Invading Nature,
6, 129-156.
Breton
G., 2005,
L'amphipode Caprella mutica Schurin, 1935, espèce
introduite et invasive, abonde dans les bassins à flot du port du
Havre (Manche orientale, France), Bulletin Trimestriel de la Société
Géologique de Normandie et des Amis du Muséum du Havre
, 91(2),
2004 (2005), 77.
Cook E.J., Jahnke M., Kerckhof F., Minchin D., Faasse M., Boos K., Ashton G., 2007, European expansion of the introduced amphipod Caprella mutica Schurin 1935, Aquatic Invasions, 2, 411-421
Schurin A., 1935, Zur Fauna der Caprelliden der Bucht Peters des
Grossen (Japanisches Meer)
, Zool. Anz., 122,
198–203.
Turcotte C., Sainte-Marie B., 2009, Synthèse de la biologie
de la caprelle japonaise (Caprella mutica),
Rapp. manus. can. sci. halieut. aquat., 2903, 28p.
La page de Caprella mutica sur le site de l'Inventaire National du Patrimoine Naturel : INPN
Les textes et images sont sous licence et ne sont pas libres de droit.
Pour les ayants-droits, connectez-vous.
Pour toute demande d'utilisation (exemple d'un formateur Bio de la FFESSM...) contactez nous ici.













")

